

Endoplasmic reticulum stress pathway PERK-eIF2α confers radioresistance in oropharyngeal carcinoma by activating NF-κB
- PMID: 28418119
- PMCID: PMC5497722
- DOI: 10.1111/cas.13260
Endoplasmic reticulum stress pathway PERK-eIF2α confers radioresistance in oropharyngeal carcinoma by activating NF-κB
Abstract
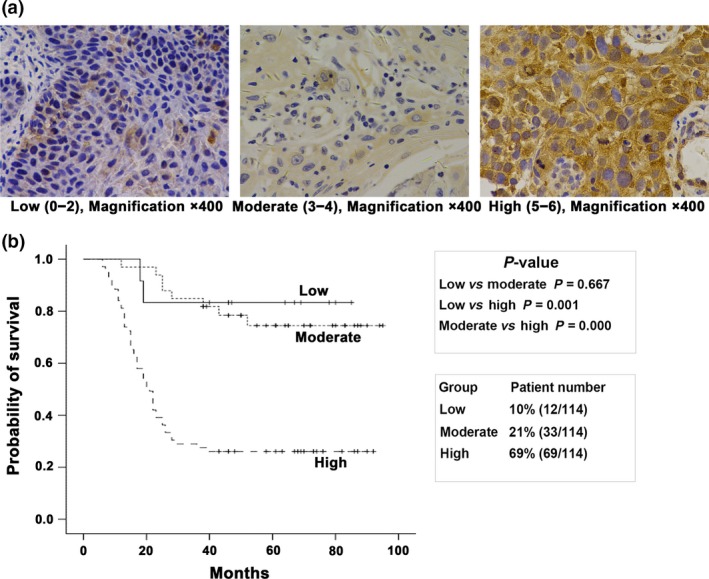
Endoplasmic reticulum stress (ERS) plays an important role in the pathogenesis and development of malignant tumors, as well as in the regulation of radiochemoresistance and chemoresistance in many malignancies. ERS signaling pathway protein kinase RNA-like endoplasmic reticulum kinase (PERK)-eukaryotic initiation factor-2 (eIF2α) may induce aberrant activation of nuclear factor-κB (NF-κB). Our previous study showed that NF-κB conferred radioresistance in lymphoma cells. However, whether PERK-eIF2α regulates radioresistance in oropharyngeal carcinoma through NF-κB activation is unknown. Herein, we showed that PERK overexpression correlated with a poor prognosis for patients with oropharyngeal carcinoma (P < 0.01). Meanwhile, the percentage of the high expression level of PERK in oropharyngeal carcinoma patients resistant to radiation was higher than in patients sensitive to radiation (77.7 and 33.3%, respectively; P < 0.05). Silencing PERK and eIF2α increased the radiosensitivity in oropharyngeal carcinoma cells and increased radiation-induced apoptosis and G2/M phase arrest. PERK-eIF2α silencing also inhibited radiation-induced NF-κB phosphorylation and increased the DNA double strand break-related proteins ATM phosphorylation. NF-κB activator TNF-α and the ATM inhibitor Ku55933 offset the regulatory effect of eIF2α on the expression of radiation-induced cell apoptosis-related proteins and the G2/M phase arrest-related proteins. These data indicate that PERK regulates radioresistance in oropharyngeal carcinoma through NF-kB activation-mediated phosphorylation of eIF2α, enhancing X-ray-induced activation of DNA DSB repair, cell apoptosis inhibition and G2/M cell cycle arrest.
Keywords: PERK; NF-κB; eIF2α; oropharyngeal carcinoma; radiotherapy.
© 2017 The Authors. Cancer Science published by John Wiley & Sons Australia, Ltd on behalf of Japanese Cancer Association.
Figures





References
-
- Jemal A, Siegel R, Ward E et al Cancer statistics, 2008. CA Cancer J Clin 2008; 58: 71–96. - PubMed
-
- Harding HP, Zhang Y, Bertolotti A, Zeng H, Ron D. Perk is essential for translational regulation and cell survival during the unfolded protein response. Mol Cell 2000; 5: 897–904. - PubMed
-
- Ma Y, Hendershot LM. The role of the unfolded protein response in tumour development: friend or foe? Nat Rev Cancer 2004; 4: 966–77. - PubMed
-
- Qi L, Wu P, Zhang X et al Inhibiting ERp29 expression enhances radiosensitivity in human nasopharyngeal carcinoma cell lines. Med Oncol 2012; 29: 721–8. - PubMed
-
- Nagelkerke A, Bussink J, van der Kogel AJ, Sweep FC, Span PN. The PERK/ATF4/LAMP3‐arm of the unfolded protein response affects radioresistance by interfering with the DNA damage response. Radiother Oncol 2013; 108: 415–21. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
