

Microhomology-mediated end joining induces hypermutagenesis at breakpoint junctions
- PMID: 28419093
- PMCID: PMC5413072
- DOI: 10.1371/journal.pgen.1006714
Microhomology-mediated end joining induces hypermutagenesis at breakpoint junctions
Abstract
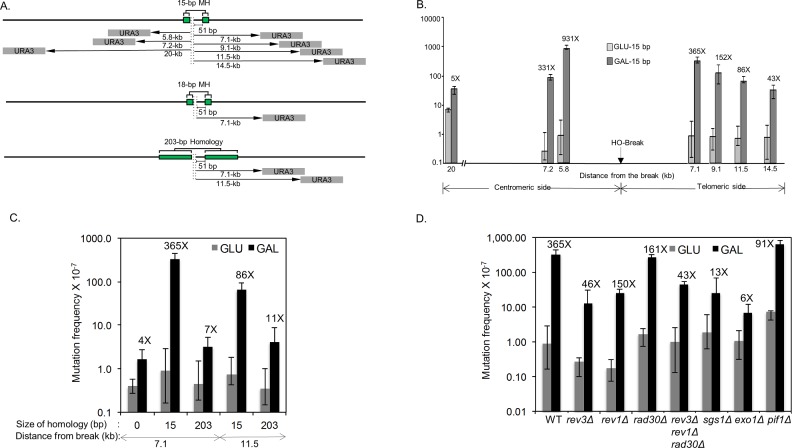
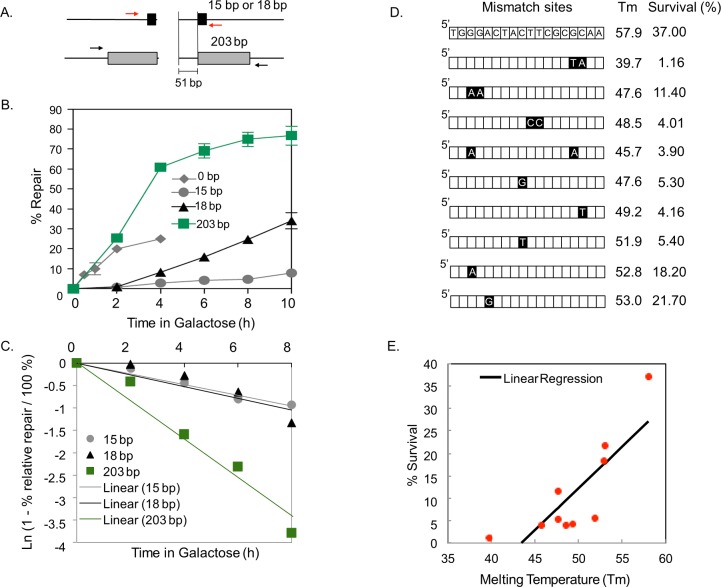
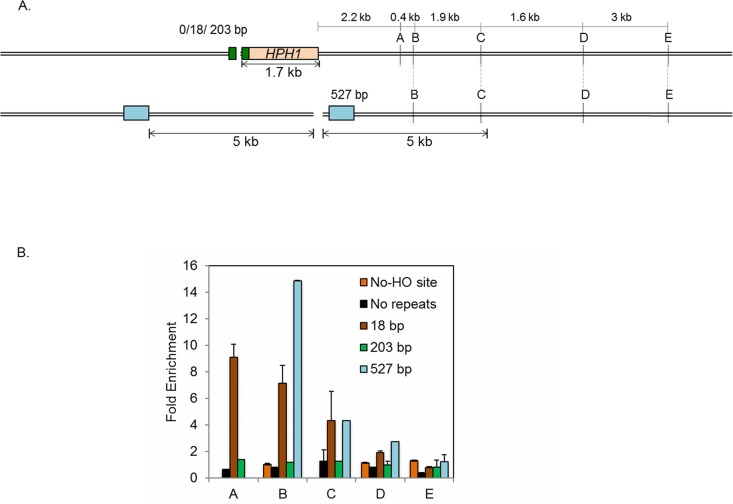
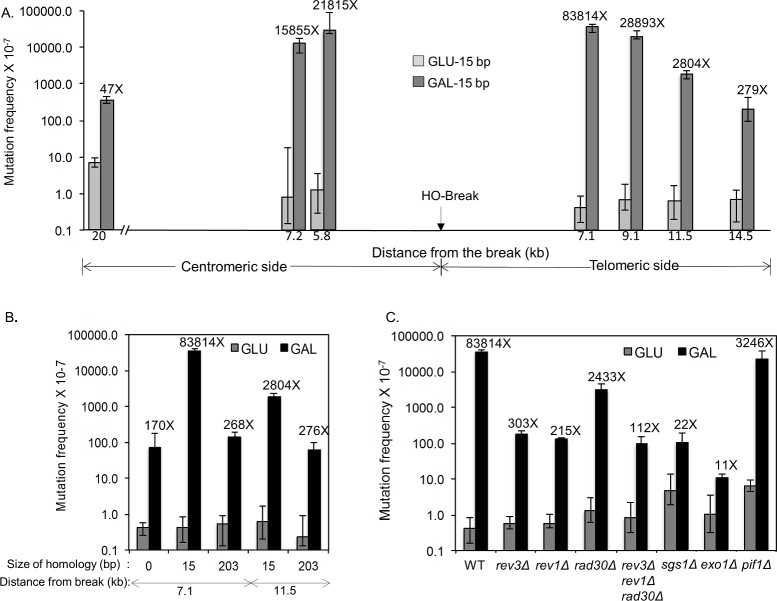
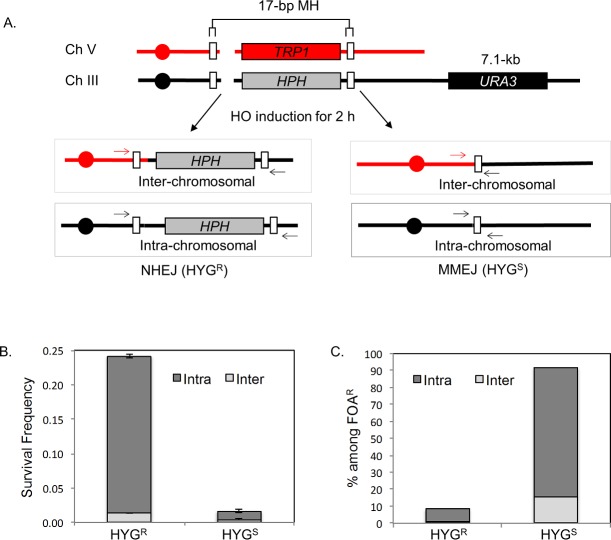
Microhomology (MH) flanking a DNA double-strand break (DSB) drives chromosomal rearrangements but its role in mutagenesis has not yet been analyzed. Here we determined the mutation frequency of a URA3 reporter gene placed at multiple locations distal to a DSB, which is flanked by different sizes (15-, 18-, or 203-bp) of direct repeat sequences for efficient repair in budding yeast. Induction of a DSB accumulates mutations in the reporter gene situated up to 14-kb distal to the 15-bp MH, but more modestly to those carrying 18- and 203-bp or no homology. Increased mutagenesis in MH-mediated end joining (MMEJ) appears coupled to its slower repair kinetics and the extensive resection occurring at flanking DNA. Chromosomal translocations via MMEJ also elevate mutagenesis of the flanking DNA sequences 7.1 kb distal to the breakpoint junction as compared to those without MH. The results suggest that MMEJ could destabilize genomes by triggering structural alterations and increasing mutation burden.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- McVey M, Lee SE. MMEJ repair of double-strand breaks (director's cut): deleted sequences and alternative endings. Trends Genet. 2008;24(11):529–38. Epub 2008/09/24. doi: 10.1016/j.tig.2008.08.007 - DOI - PMC - PubMed
-
- Sfeir A, Symington LS. Microhomology-Mediated End Joining: A Back-up Survival Mechanism or Dedicated Pathway? Trends Biochem Sci. 2015;40(11):701–14. Epub 2015/10/07. PubMed Central PMCID: PMCPMC4638128. doi: 10.1016/j.tibs.2015.08.006 - DOI - PMC - PubMed
-
- Lieber MR, Lu H, Gu J, Schwarz K. Flexibility in the order of action and in the enzymology of the nuclease, polymerases, and ligase of vertebrate non-homologous DNA end joining: relevance to cancer, aging, and the immune system. Cell Res. 2008;18(1):125–33. Epub 2007/12/19. doi: 10.1038/cr.2007.108 - DOI - PubMed
-
- Lieber MR. Mechanisms of human lymphoid chromosomal translocations. Nat Rev Cancer. 2016;16(6):387–98. Epub 2016/05/26. doi: 10.1038/nrc.2016.40 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
