

Melatonin, a Full Service Anti-Cancer Agent: Inhibition of Initiation, Progression and Metastasis
- PMID: 28420185
- PMCID: PMC5412427
- DOI: 10.3390/ijms18040843
Melatonin, a Full Service Anti-Cancer Agent: Inhibition of Initiation, Progression and Metastasis
Abstract
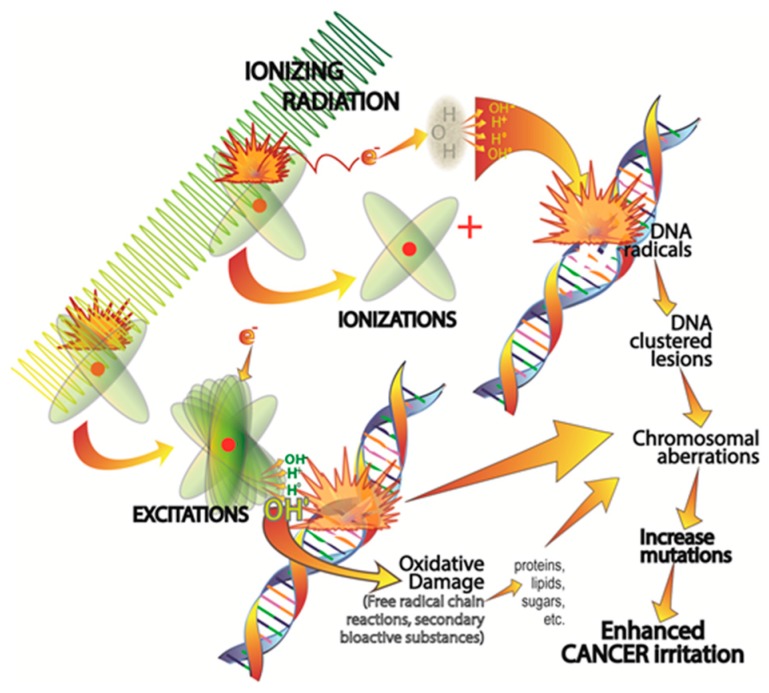
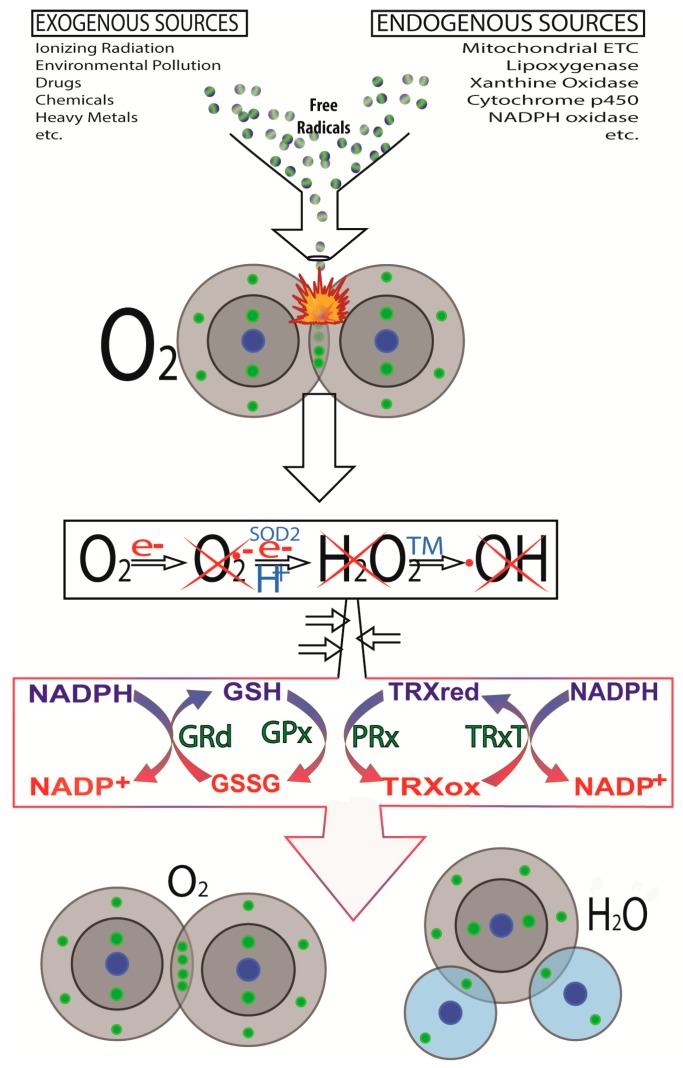
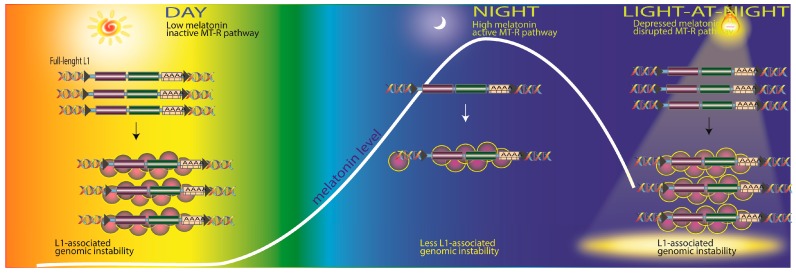
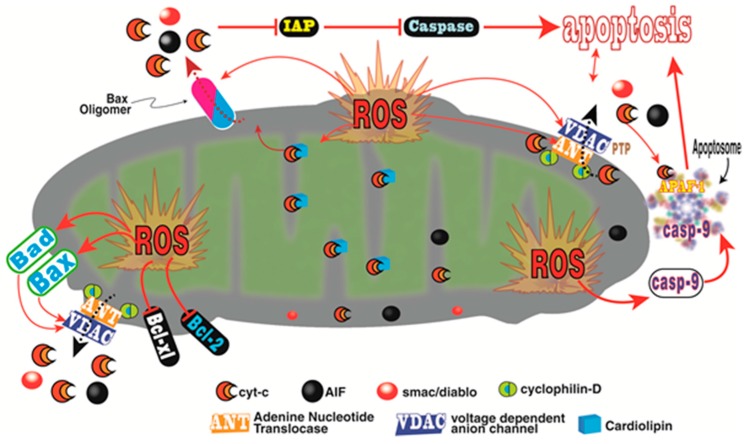
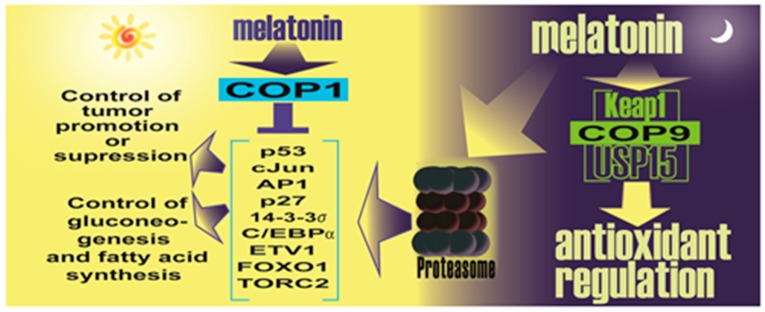
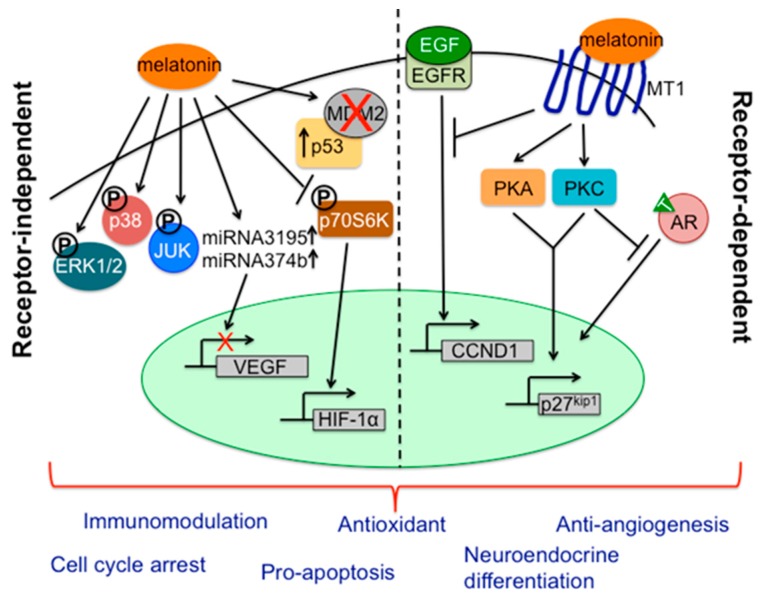
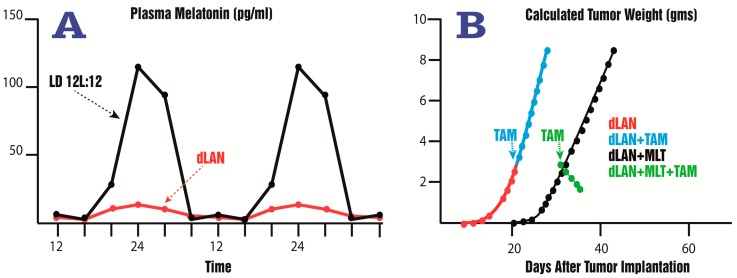
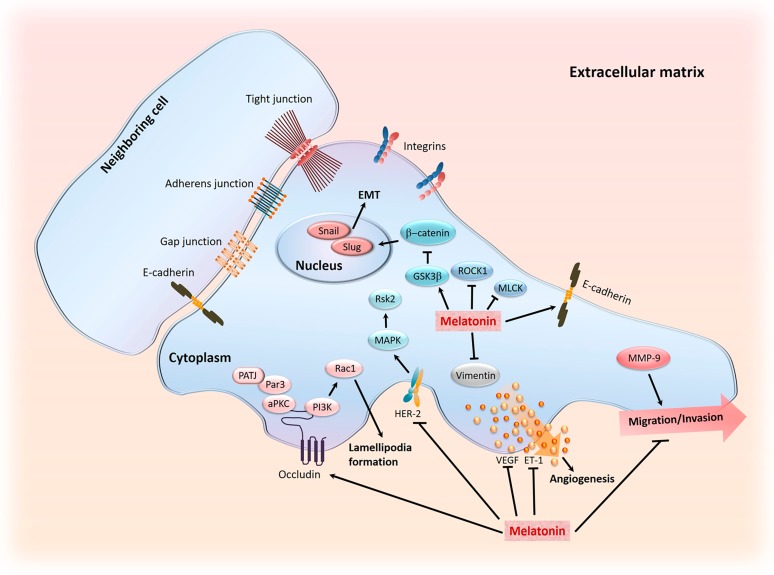
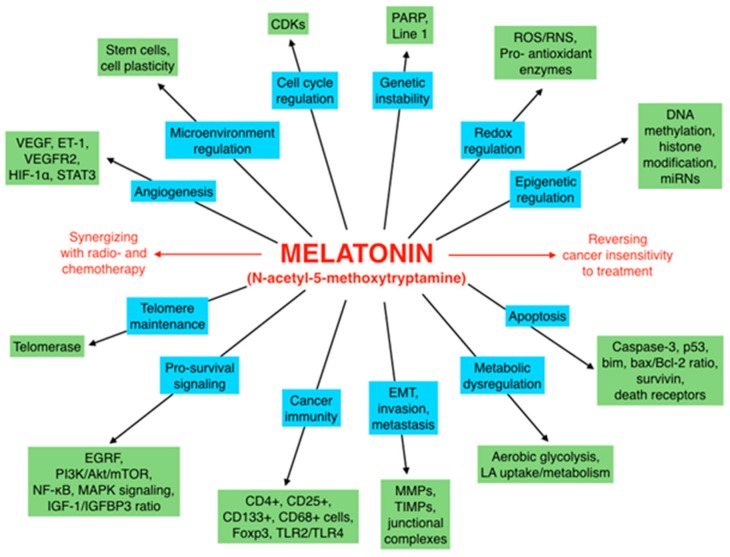
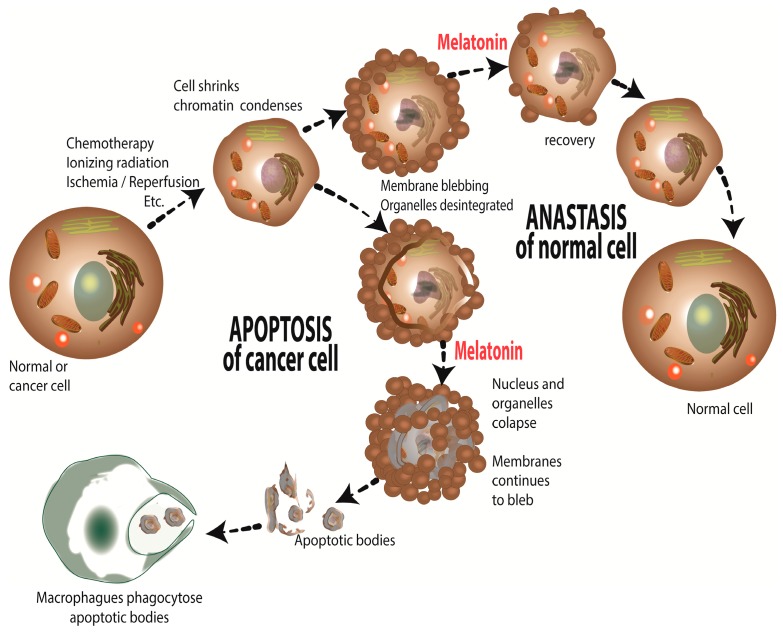
There is highly credible evidence that melatonin mitigates cancer at the initiation, progression and metastasis phases. In many cases, the molecular mechanisms underpinning these inhibitory actions have been proposed. What is rather perplexing, however, is the large number of processes by which melatonin reportedly restrains cancer development and growth. These diverse actions suggest that what is being observed are merely epiphenomena of an underlying more fundamental action of melatonin that remains to be disclosed. Some of the arresting actions of melatonin on cancer are clearly membrane receptor-mediated while others are membrane receptor-independent and involve direct intracellular actions of this ubiquitously-distributed molecule. While the emphasis of melatonin/cancer research has been on the role of the indoleamine in restraining breast cancer, this is changing quickly with many cancer types having been shown to be susceptible to inhibition by melatonin. There are several facets of this research which could have immediate applications at the clinical level. Many studies have shown that melatonin's co-administration improves the sensitivity of cancers to inhibition by conventional drugs. Even more important are the findings that melatonin renders cancers previously totally resistant to treatment sensitive to these same therapies. Melatonin also inhibits molecular processes associated with metastasis by limiting the entrance of cancer cells into the vascular system and preventing them from establishing secondary growths at distant sites. This is of particular importance since cancer metastasis often significantly contributes to death of the patient. Another area that deserves additional consideration is related to the capacity of melatonin in reducing the toxic consequences of anti-cancer drugs while increasing their efficacy. Although this information has been available for more than a decade, it has not been adequately exploited at the clinical level. Even if the only beneficial actions of melatonin in cancer patients are its ability to attenuate acute and long-term drug toxicity, melatonin should be used to improve the physical wellbeing of the patients. The experimental findings, however, suggest that the advantages of using melatonin as a co-treatment with conventional cancer therapies would far exceed improvements in the wellbeing of the patients.
Keywords: angiogenesis; antioxidant; apoptosis; breast; chemotherapy; free radicals; invasion; ionizing radiation; melatonin receptors; molecular mechanisms; prostate.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Tan D.X., Chen L.D., Poeggeler B., Manchester L.C., Reiter R.J. Melatonin: A potent, endogenous hydroxyl radical scavenger. J. Pineal Res. 1993;1:57–60. - PubMed
-
- Poeggeler B., Saarela S., Reiter R.J., Tan D.X., Chen L.D., Manchester L.C., Barlow-Walden L. Melatonin—A highly potent endogenous radical scavenger and electron donor: New aspects of the oxidation chemistry of the indole assessed in vitro. Ann. N. Y. Acad. Sci. 1994;738:419–420. doi: 10.1111/j.1749-6632.1994.tb21831.x. - DOI - PubMed
-
- Zavodnik I.B., Domanski A.V., Lapshina E.A., Bryszewska M., Reiter R.J. Melatonin directly scavenges free radicals generated in red blood cells and in a cell-free system: Chemiluminescence measurements and theoretical calculations. Life Sci. 2006;79:391–400. doi: 10.1016/j.lfs.2006.01.030. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
