

Synergistic Internal Ribosome Entry Site/MicroRNA-Based Approach for Flavivirus Attenuation and Live Vaccine Development
- PMID: 28420742
- PMCID: PMC5395672
- DOI: 10.1128/mBio.02326-16
Synergistic Internal Ribosome Entry Site/MicroRNA-Based Approach for Flavivirus Attenuation and Live Vaccine Development
Abstract
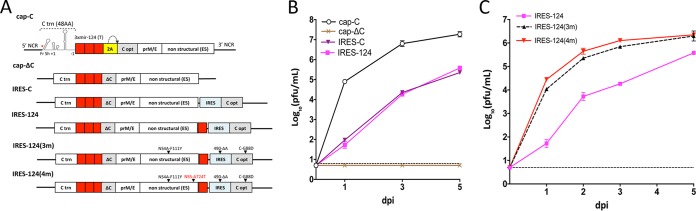
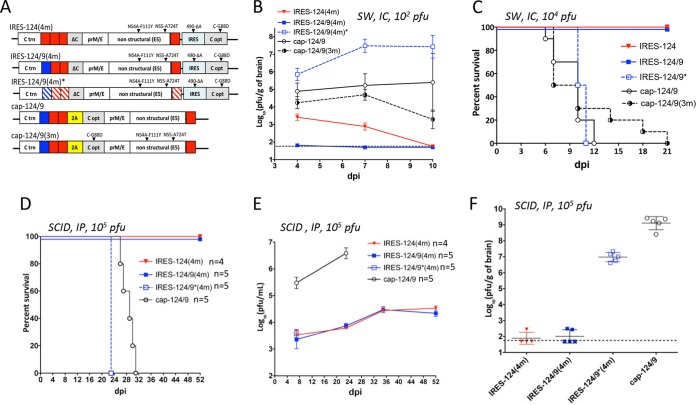
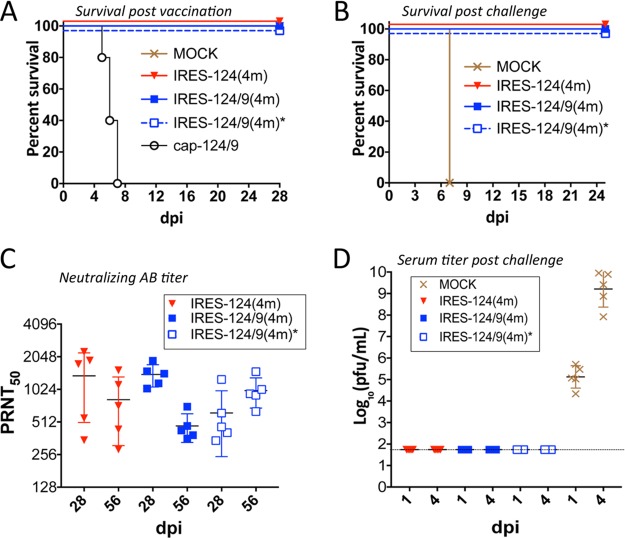
The recent emergence of Zika virus underscores the need for new strategies for a rapid development of safe flavivirus vaccines. Using another flavivirus (Langat virus [LGTV]) that belongs to the group of tick-borne flaviviruses as a model, we describe a dual strategy for virus attenuation which synergistically accesses the specificity of microRNA (miRNA) genome targeting and the effectiveness of internal ribosome entry site (IRES) insertion. To increase the stability and immunogenicity of bicistronic LGTVs, we developed a novel approach in which the capsid (C) protein gene was relocated into the 3' noncoding region (NCR) and expressed under translational control from an IRES. Engineered bicistronic LGTVs carrying multiple target sequences for brain-specific miRNAs were stable in Vero cells and induced adaptive immunity in mice. Importantly, miRNA-targeted bicistronic LGTVs were not pathogenic for either newborn mice after intracranial inoculation or adult immunocompromised mice (SCID or type I interferon receptor knockout) after intraperitoneal injection. Moreover, bicistronic LGTVs were restricted for replication in tick-derived cells, suggesting an interruption of viral transmission in nature by arthropod vectors. This approach is suitable for reliable attenuation of many flaviviruses and may enable development of live attenuated flavivirus vaccines.IMPORTANCE The recent emergence of Zika virus underscores the need for new strategies for a rapid development of safe flavivirus vaccines. Allied separately attenuating approaches based on (i) microRNA genome targeting and (ii) internal ribosome entry site insertion are not sufficient for relievable attenuation of neurotropic flavivirus pathogenesis. Here, we describe a novel dual strategy that combines the specificity of miRNA-based and the effectiveness of IRES-based attenuating approaches, allowing us to overcome these critical limitations. This developed approach provides a robust platform for reliable attenuation of many flaviviruses and may enable development of live flavivirus vaccines.
Keywords: attenuation; flavivirus; internal ribosome entry site; microRNA; vaccine.
Copyright © 2017 Tsetsarkin et al.
Figures
Similar articles
-
Stable and Highly Immunogenic MicroRNA-Targeted Single-Dose Live Attenuated Vaccine Candidate against Tick-Borne Encephalitis Constructed Using Genetic Backbone of Langat Virus.mBio. 2019 Apr 23;10(2):e02904-18. doi: 10.1128/mBio.02904-18. mBio. 2019. PMID: 31015334 Free PMC article.
-
Kissing-loop interaction between 5' and 3' ends of tick-borne Langat virus genome 'bridges the gap' between mosquito- and tick-borne flaviviruses in mechanisms of viral RNA cyclization: applications for virus attenuation and vaccine development.Nucleic Acids Res. 2016 Apr 20;44(7):3330-50. doi: 10.1093/nar/gkw061. Epub 2016 Feb 4. Nucleic Acids Res. 2016. PMID: 26850640 Free PMC article.
-
Tick-borne Langat/mosquito-borne dengue flavivirus chimera, a candidate live attenuated vaccine for protection against disease caused by members of the tick-borne encephalitis virus complex: evaluation in rhesus monkeys and in mosquitoes.J Virol. 2001 Sep;75(17):8259-67. doi: 10.1128/jvi.75.17.8259-8267.2001. J Virol. 2001. PMID: 11483771 Free PMC article.
-
Chimeric flaviviruses: novel vaccines against dengue fever, tick-borne encephalitis, and Japanese encephalitis.Adv Virus Res. 2003;61:469-509. doi: 10.1016/s0065-3527(03)61013-4. Adv Virus Res. 2003. PMID: 14714441 Review.
-
Identifying Attenuating Mutations: Tools for a New Vaccine Design against Flaviviruses.Intervirology. 2017;60(1-2):8-18. doi: 10.1159/000479966. Epub 2017 Sep 5. Intervirology. 2017. PMID: 28869941 Review.
Cited by
-
Stable and Highly Immunogenic MicroRNA-Targeted Single-Dose Live Attenuated Vaccine Candidate against Tick-Borne Encephalitis Constructed Using Genetic Backbone of Langat Virus.mBio. 2019 Apr 23;10(2):e02904-18. doi: 10.1128/mBio.02904-18. mBio. 2019. PMID: 31015334 Free PMC article.
-
Development of a Bicistronic Yellow Fever Live Attenuated Vaccine with Reduced Neurovirulence and Viscerotropism.Microbiol Spectr. 2022 Oct 26;10(5):e0224622. doi: 10.1128/spectrum.02246-22. Epub 2022 Aug 18. Microbiol Spectr. 2022. PMID: 35980184 Free PMC article.
-
Interactions of host miRNAs in the flavivirus 3´UTR genome: From bioinformatics predictions to practical approaches.Front Cell Infect Microbiol. 2022 Oct 13;12:976843. doi: 10.3389/fcimb.2022.976843. eCollection 2022. Front Cell Infect Microbiol. 2022. PMID: 36310869 Free PMC article. Review.
-
Novel Approach for Insertion of Heterologous Sequences into Full-Length ZIKV Genome Results in Superior Level of Gene Expression and Insert Stability.Viruses. 2020 Jan 3;12(1):61. doi: 10.3390/v12010061. Viruses. 2020. PMID: 31947825 Free PMC article.
-
A Roadmap for Tick-Borne Flavivirus Research in the "Omics" Era.Front Cell Infect Microbiol. 2017 Dec 22;7:519. doi: 10.3389/fcimb.2017.00519. eCollection 2017. Front Cell Infect Microbiol. 2017. PMID: 29312896 Free PMC article. Review.
References
-
- Tsetsarkin KA, Liu G, Shen K, Pletnev AG. 2016. Kissing-loop interaction between 5′ and 3′ ends of tick-borne Langat virus genome “bridges the gap” between mosquito- and tick-borne flaviviruses in mechanisms of viral RNA cyclization: applications for virus attenuation and vaccine development. Nucleic Acids Res 44:3330–3350. doi:10.1093/nar/gkw061. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
