

A critical-like collective state leads to long-range cell communication in Dictyostelium discoideum aggregation
- PMID: 28422986
- PMCID: PMC5396852
- DOI: 10.1371/journal.pbio.1002602
A critical-like collective state leads to long-range cell communication in Dictyostelium discoideum aggregation
Abstract
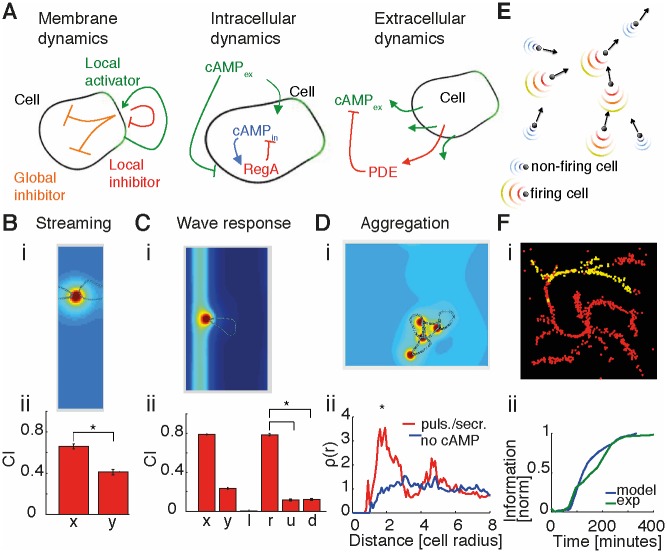
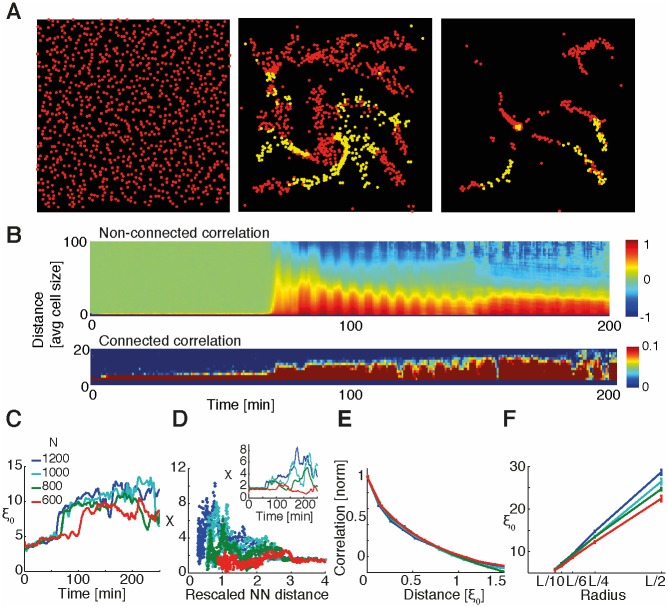
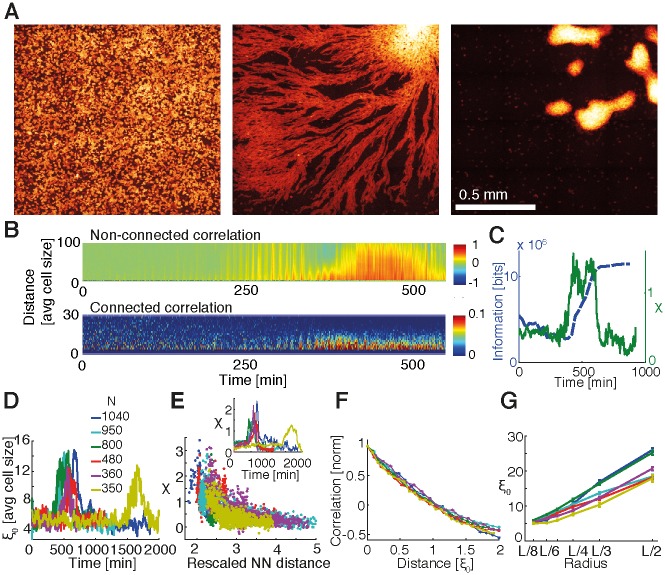
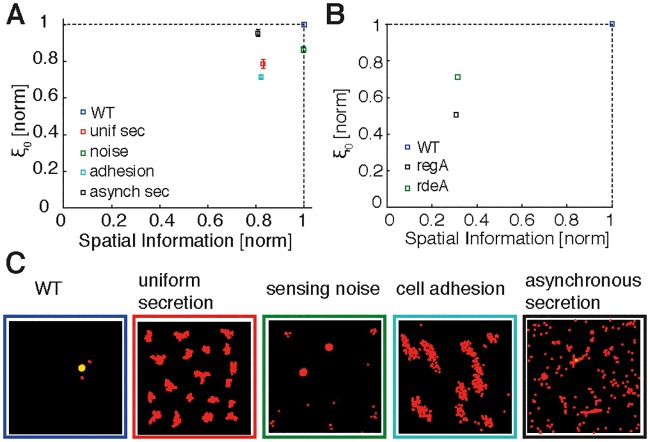
The transition from single-cell to multicellular behavior is important in early development but rarely studied. The starvation-induced aggregation of the social amoeba Dictyostelium discoideum into a multicellular slug is known to result from single-cell chemotaxis towards emitted pulses of cyclic adenosine monophosphate (cAMP). However, how exactly do transient, short-range chemical gradients lead to coherent collective movement at a macroscopic scale? Here, we developed a multiscale model verified by quantitative microscopy to describe behaviors ranging widely from chemotaxis and excitability of individual cells to aggregation of thousands of cells. To better understand the mechanism of long-range cell-cell communication and hence aggregation, we analyzed cell-cell correlations, showing evidence of self-organization at the onset of aggregation (as opposed to following a leader cell). Surprisingly, cell collectives, despite their finite size, show features of criticality known from phase transitions in physical systems. By comparing wild-type and mutant cells with impaired aggregation, we found the longest cell-cell communication distance in wild-type cells, suggesting that criticality provides an adaptive advantage and optimally sized aggregates for the dispersal of spores.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Ion Signaling in Cell Motility and Development in Dictyostelium discoideum.Biomolecules. 2024 Jul 10;14(7):830. doi: 10.3390/biom14070830. Biomolecules. 2024. PMID: 39062545 Free PMC article. Review.
-
A Mutant of Dictyostelium discoideum, KI-Cell, as a Model of Collective Cell Migration Independent of Chemotaxis.Methods Mol Biol. 2024;2828:37-43. doi: 10.1007/978-1-0716-4023-4_4. Methods Mol Biol. 2024. PMID: 39147968
-
Diverse chemotactic responses of Dictyostelium discoideum amoebae in the developing (temporal) and stationary (spatial) concentration gradients of folic acid, cAMP, Ca(2+) and Mg(2+).Cell Motil Cytoskeleton. 2002 Sep;53(1):1-25. doi: 10.1002/cm.10052. Cell Motil Cytoskeleton. 2002. PMID: 12211112
-
Continuous emergence of phototaxis in Dictyostelium discoideum.PLoS One. 2025 May 19;20(5):e0321614. doi: 10.1371/journal.pone.0321614. eCollection 2025. PLoS One. 2025. PMID: 40388399 Free PMC article.
-
Follow the leader.Dev Cell. 2003 Mar;4(3):291-3. doi: 10.1016/s1534-5807(03)00064-9. Dev Cell. 2003. PMID: 12636911 Review.
Cited by
-
Exploring the criticality hypothesis using programmable swarm robots with Vicsek-like interactions.J R Soc Interface. 2023 Jul;20(204):20230176. doi: 10.1098/rsif.2023.0176. Epub 2023 Jul 19. J R Soc Interface. 2023. PMID: 37464802 Free PMC article.
-
Matrix feedback enables diverse higher-order patterning of the extracellular matrix.PLoS Comput Biol. 2019 Oct 28;15(10):e1007251. doi: 10.1371/journal.pcbi.1007251. eCollection 2019 Oct. PLoS Comput Biol. 2019. PMID: 31658254 Free PMC article.
-
Adaptive self-organization in the embryo: its importance to adult anatomy and to tissue engineering.J Anat. 2018 Apr;232(4):524-533. doi: 10.1111/joa.12691. Epub 2017 Oct 10. J Anat. 2018. PMID: 29023694 Free PMC article. Review.
-
Bacterial surface motility is modulated by colony-scale flow and granular jamming.J R Soc Interface. 2020 Jun;17(167):20200147. doi: 10.1098/rsif.2020.0147. Epub 2020 Jun 24. J R Soc Interface. 2020. PMID: 32574537 Free PMC article.
-
The hallmarks of living systems: towards creating artificial cells.Interface Focus. 2018 Oct 6;8(5):20180023. doi: 10.1098/rsfs.2018.0023. Epub 2018 Aug 17. Interface Focus. 2018. PMID: 30443324 Free PMC article. Review.
References
-
- Vicsek T, Zafeiris A. Collective motion. Physics Reports. 2012;517(3–4):71–140. 10.1016/j.physrep.2012.03.004. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
