

The glycogen synthase kinase MoGsk1, regulated by Mps1 MAP kinase, is required for fungal development and pathogenicity in Magnaporthe oryzae
- PMID: 28424497
- PMCID: PMC5430414
- DOI: 10.1038/s41598-017-01006-w
The glycogen synthase kinase MoGsk1, regulated by Mps1 MAP kinase, is required for fungal development and pathogenicity in Magnaporthe oryzae
Erratum in
-
Publisher Correction: The glycogen synthase kinase MoGsk1, regulated by Mps1 MAP kinase, is required for fungal development and pathogenicity in Magnaporthe oryzae.Sci Rep. 2020 Jan 21;10(1):1187. doi: 10.1038/s41598-020-58148-7. Sci Rep. 2020. PMID: 31959847 Free PMC article.
Abstract
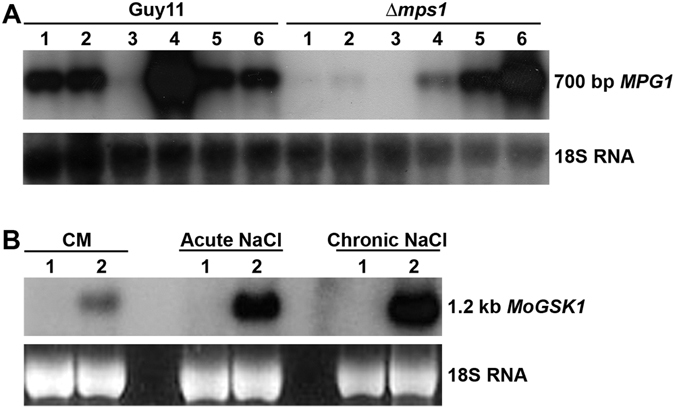
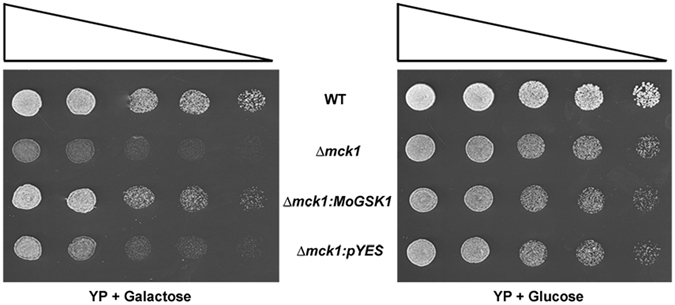
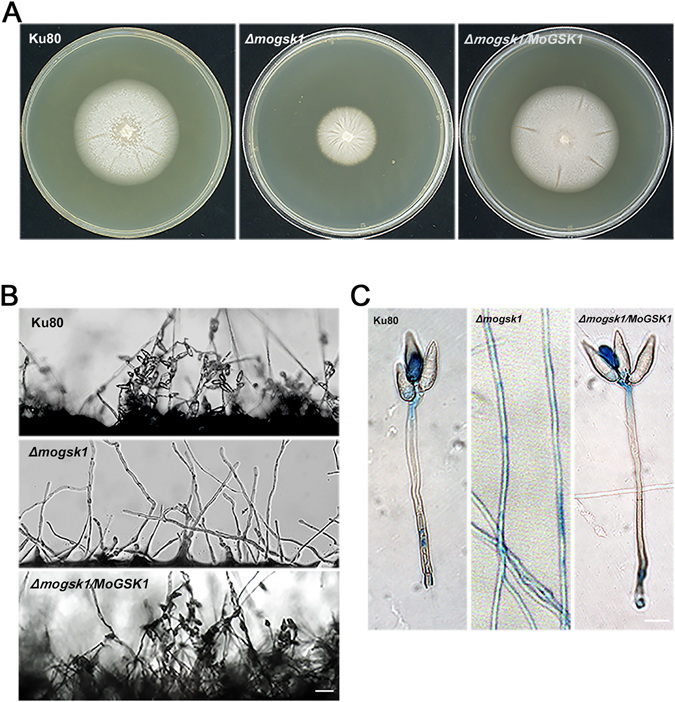
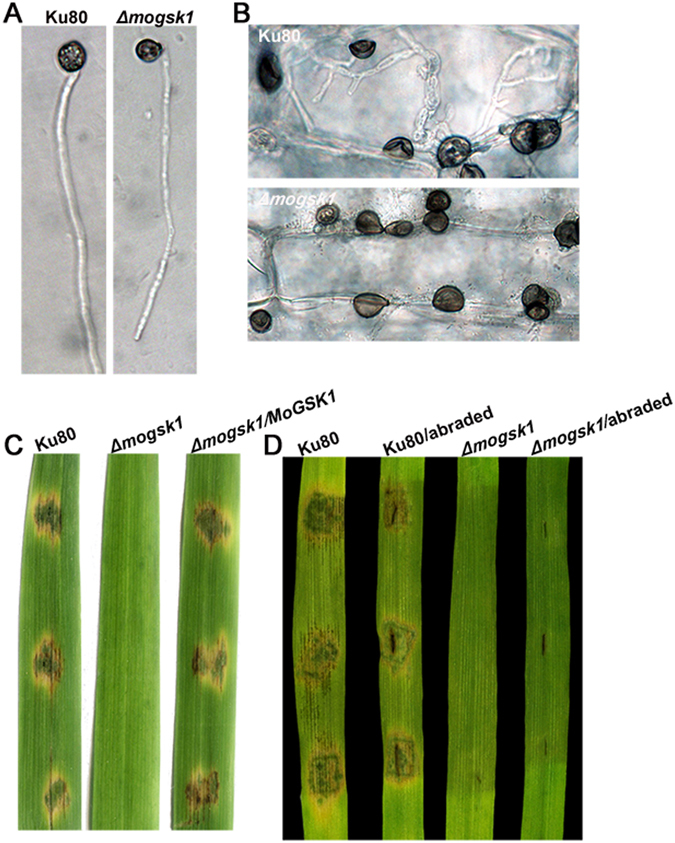
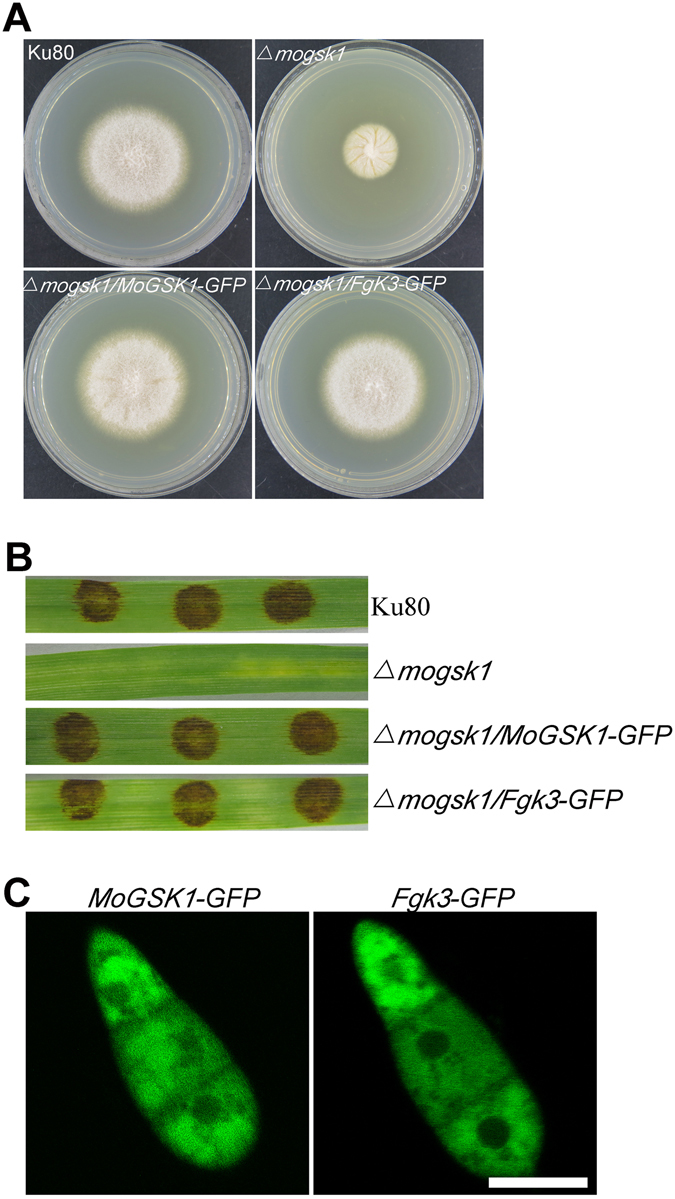
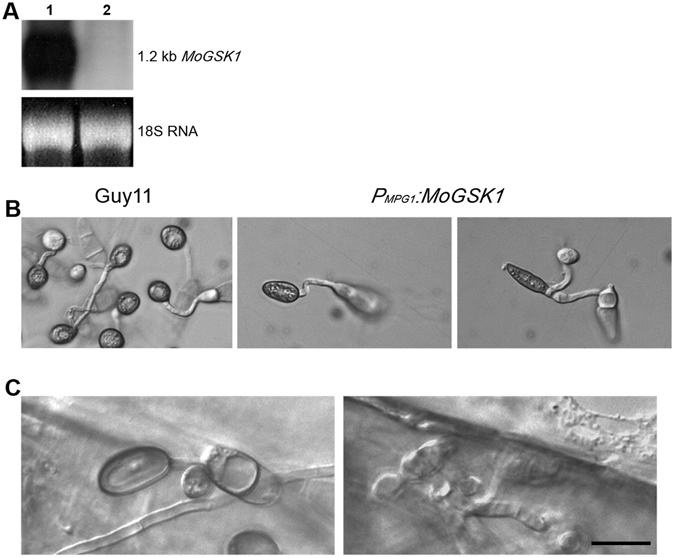
Magnaporthe oryzae, the causal agent of blast disease, is one of the most destructive plant pathogens, causing significant yield losses on staple crops such as rice and wheat. The fungus infects plants with a specialized cell called an appressorium, whose development is tightly regulated by MAPK signaling pathways following the activation of upstream sensors in response to environmental stimuli. Here, we show the expression of the Glycogen synthase kinase 3 (GSK3) MoGSK1 in M. oryzae is regulated by Mps1 MAP kinase, particularly under the stressed conditions. Thus, MoGSK1 is functionally characterized in this study. MoGsk1 is functionally homologues to the Saccharomyces cerevisiae GSK3 homolog MCK1. Gene replacement of MoGSK1 caused significant delay in mycelial growth, complete loss of conidiation and inability to penetrate the host surface by mycelia-formed appressorium-like structures, consequently resulting in loss of pathogenicity. However, the developmental and pathogenic defects of Δmogsk1 are recovered via the heterologous expression of Fusarium graminearum GSK3 homolog gene FGK3, whose coding products also shows the similar cytoplasmic localization as MoGsk1 does in M. oryzae. By contrast, overexpression of MoGSK1 produced deformed appressoria in M. oryzae. In summary, our results suggest that MoGsk1, as a highly conservative signal modulator, dictates growth, conidiation and pathogenicity of M. oryzae.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Motallebi P, Javan-Nikkhah M, Okhovvat M, Berdi Fotouhifar K, Hossien Mosahebi G. Differentiation of Magnaporthe species complex by rep-PCR genomic fingerprinting. Commun. Agric. Appl. Biol. Sci. 2009;74:821–829. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
