

Interactions of the Mechanosensitive Channels with Extracellular Matrix, Integrins, and Cytoskeletal Network in Osmosensation
- PMID: 28424587
- PMCID: PMC5380722
- DOI: 10.3389/fnmol.2017.00096
Interactions of the Mechanosensitive Channels with Extracellular Matrix, Integrins, and Cytoskeletal Network in Osmosensation
Abstract
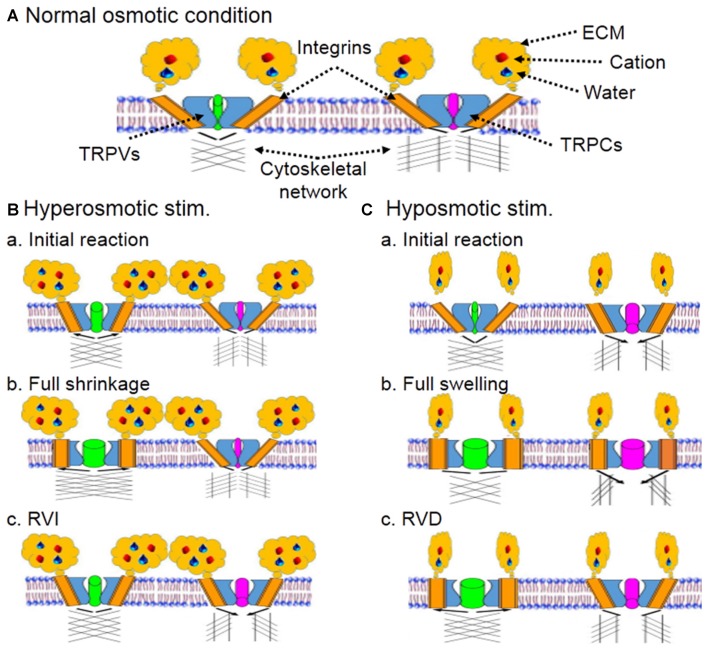
Life is maintained in a sea water-like internal environment. The homeostasis of this environment is dependent on osmosensory system translation of hydromineral information into osmotic regulatory machinery at system, tissue and cell levels. In the osmosensation, hydromineral information can be converted into cellular reactions through osmoreceptors, which changes thirst and drinking, secretion of antidiuretic vasopressin (VP), reabsorption of water and salt in the kidneys at systemic level as well as cellular metabolic activity and survival status at tissue level. The key feature of osmosensation is the activation of mechanoreceptors or mechanosensors, particularly transient receptor potential vallinoid (TRPV) and canonical (TRPC) family channels, which increases cytosolic Ca2+ levels, activates osmosensory cells including VP neurons and triggers a series of secondary reactions. TRPV channels are sensitive to both hyperosmotic and hyposmotic stimuli while TRPC channels are more sensitive to hyposmotic challenge in neurons. The activation of TRP channels relies on changes in cell volume, membrane stretch and cytoskeletal reorganization as well as hydration status of extracellular matrix (ECM) and activity of integrins. Different families of TRP channels could be activated differently in response to hyperosmotic and hyposmotic stimuli in different spatiotemporal orders, leading to differential reactions of osmosensory cells. Together, they constitute the osmosensory machinery. The activation of this osmoreceptor complex is also associated with the activity of other osmolarity-regulating organelles, such as water channel protein aquaporins, Na-K-2Cl cotransporters, volume-sensitive anion channels, sodium pump and purinergic receptors in addition to intercellular interactions, typically astrocytic neuronal interactions. In this article, we review our current understandings of the composition of osmoreceptors and the processes of osmosensation.
Keywords: cytoskeleton; extracellular matrix; integrin; transient receptor potential canonical channel; transient receptor potential vallinoid channel; vasopressin.
Figures
Similar articles
-
Astroglial Modulation of Hydromineral Balance and Cerebral Edema.Front Mol Neurosci. 2018 Jun 12;11:204. doi: 10.3389/fnmol.2018.00204. eCollection 2018. Front Mol Neurosci. 2018. PMID: 29946238 Free PMC article. Review.
-
Contribution of TRPV channels to osmosensory transduction, thirst, and vasopressin release.Kidney Int. 2008 Apr;73(7):811-5. doi: 10.1038/sj.ki.5002788. Epub 2008 Jan 16. Kidney Int. 2008. PMID: 18200003 Review.
-
Osmosensation in TRPV2 dominant negative expressing skeletal muscle fibres.J Physiol. 2015 Sep 1;593(17):3849-63. doi: 10.1113/JP270522. Epub 2015 Aug 10. J Physiol. 2015. PMID: 26108786 Free PMC article.
-
The mechanosensitive nature of TRPV channels.Pflugers Arch. 2005 Oct;451(1):193-203. doi: 10.1007/s00424-005-1424-4. Epub 2005 May 21. Pflugers Arch. 2005. PMID: 15909178 Review.
-
Osmosensation in vasopressin neurons: changing actin density to optimize function.Trends Neurosci. 2010 Feb;33(2):76-83. doi: 10.1016/j.tins.2009.11.004. Epub 2009 Dec 4. Trends Neurosci. 2010. PMID: 19963290 Review.
Cited by
-
Focusing on Mechanoregulation Axis in Fibrosis: Sensing, Transduction and Effecting.Front Mol Biosci. 2022 Mar 11;9:804680. doi: 10.3389/fmolb.2022.804680. eCollection 2022. Front Mol Biosci. 2022. PMID: 35359592 Free PMC article. Review.
-
Involvement of extrasynaptic glutamate in physiological and pathophysiological changes of neuronal excitability.Cell Mol Life Sci. 2018 Aug;75(16):2917-2949. doi: 10.1007/s00018-018-2837-5. Epub 2018 May 15. Cell Mol Life Sci. 2018. PMID: 29766217 Free PMC article. Review.
-
Therapeutic Potential of Oxytocin in Atherosclerotic Cardiovascular Disease: Mechanisms and Signaling Pathways.Front Neurosci. 2019 May 21;13:454. doi: 10.3389/fnins.2019.00454. eCollection 2019. Front Neurosci. 2019. PMID: 31178679 Free PMC article. Review.
-
TRPV1-dependent NKCC1 activation in mouse lens involves integrin and the tubulin cytoskeleton.J Cell Physiol. 2024 Nov;239(11):e31369. doi: 10.1002/jcp.31369. Epub 2024 Jul 16. J Cell Physiol. 2024. PMID: 39014912
-
Tumour Initiation: a Discussion on Evidence for a "Load-Trigger" Mechanism.Cell Biochem Biophys. 2019 Dec;77(4):293-308. doi: 10.1007/s12013-019-00888-z. Epub 2019 Oct 9. Cell Biochem Biophys. 2019. PMID: 31598831 Free PMC article. Review.
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous