

The integrity of the nucleus of the lateral olfactory tract is essential for the normal functioning of the olfactory system
- PMID: 28424894
- PMCID: PMC5676812
- DOI: 10.1007/s00429-017-1422-2
The integrity of the nucleus of the lateral olfactory tract is essential for the normal functioning of the olfactory system
Abstract
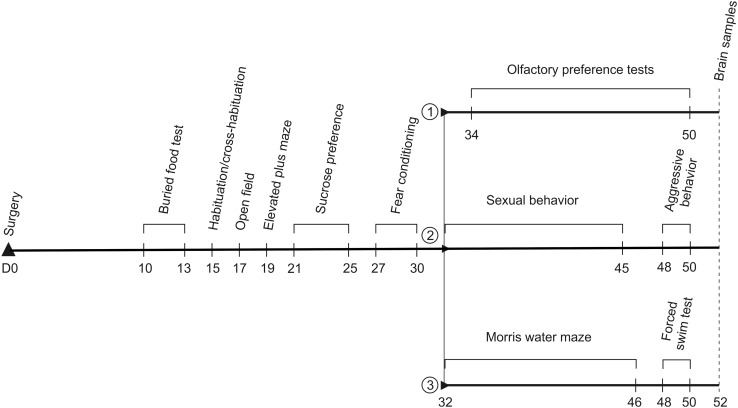
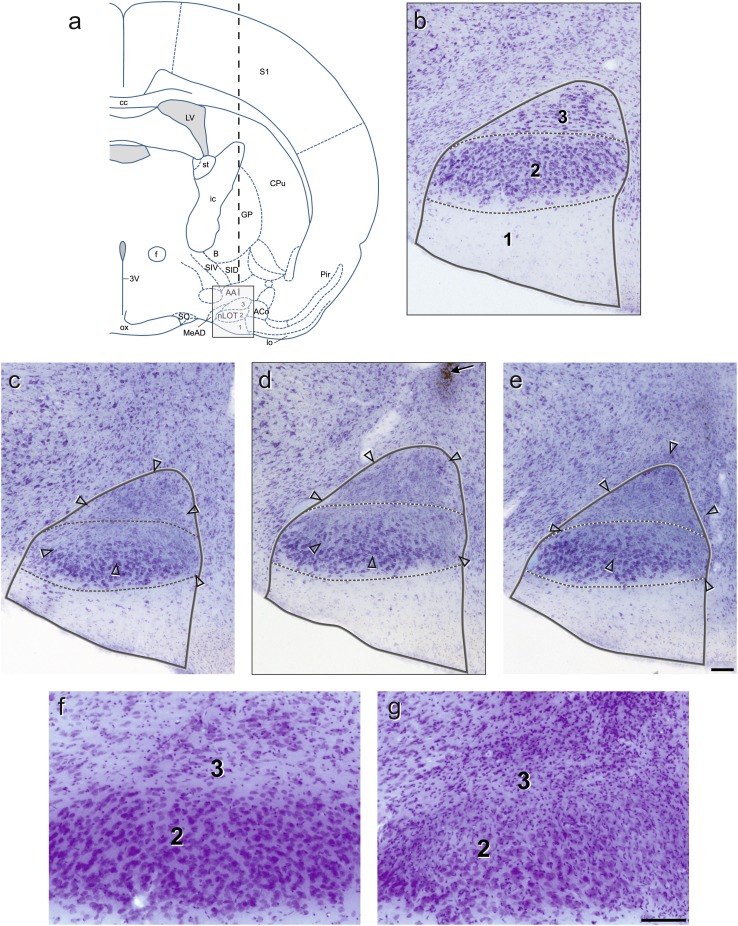
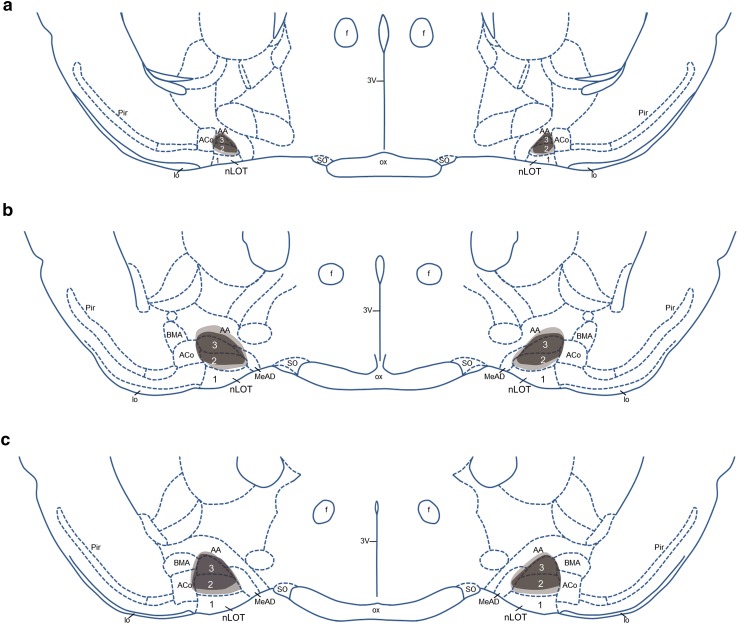
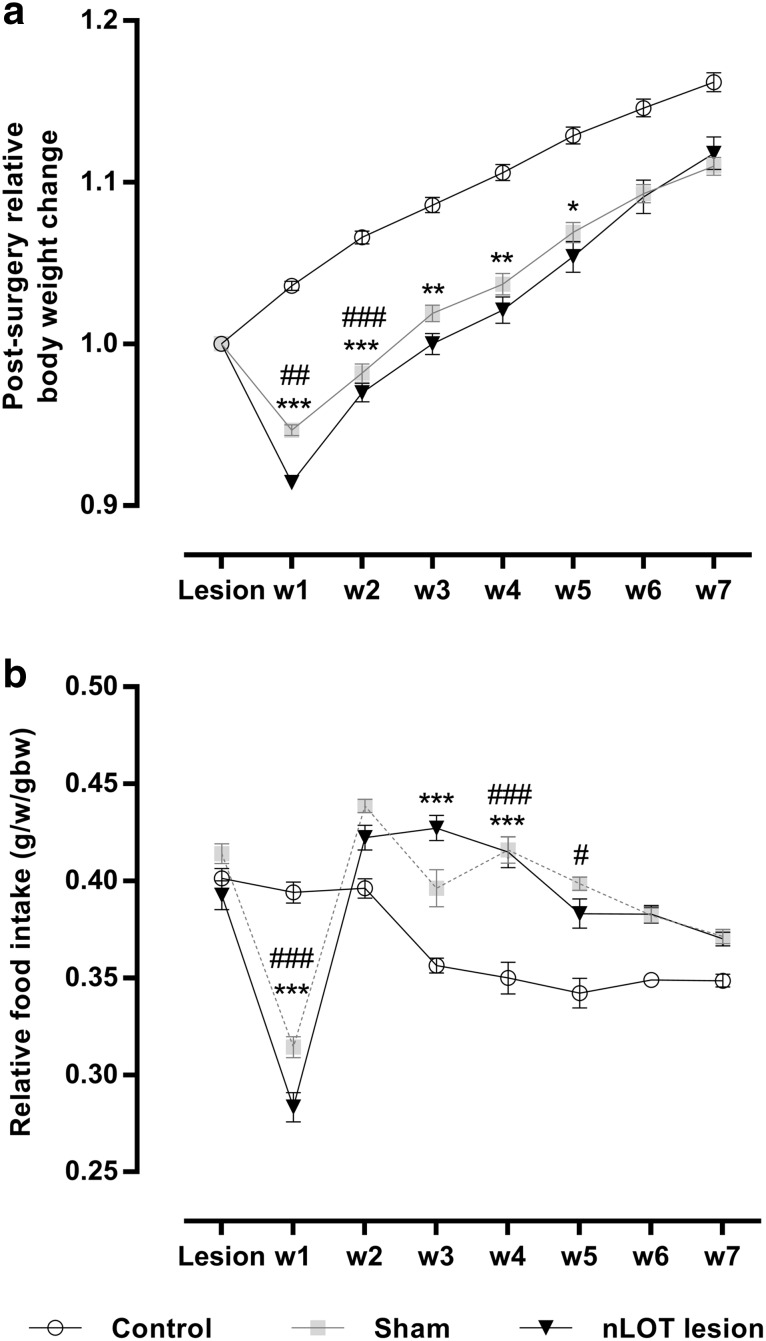
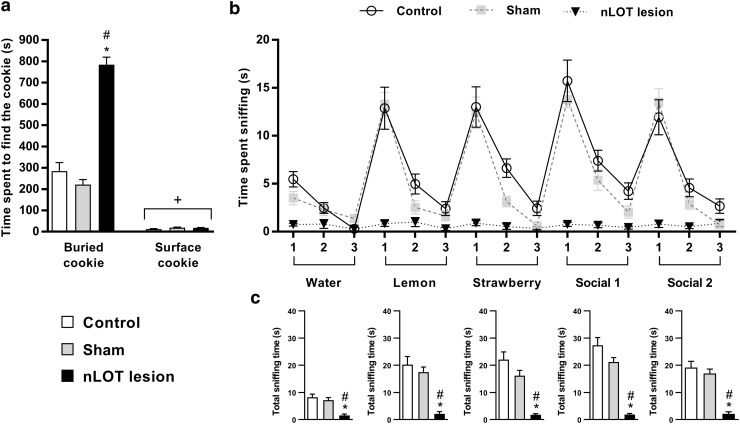
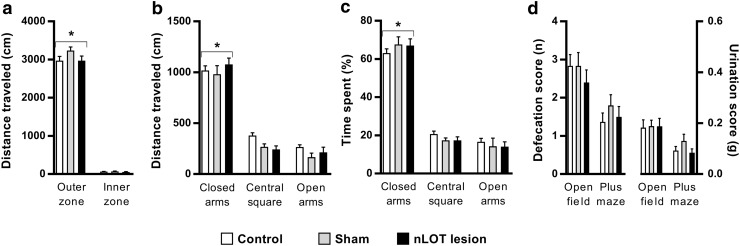
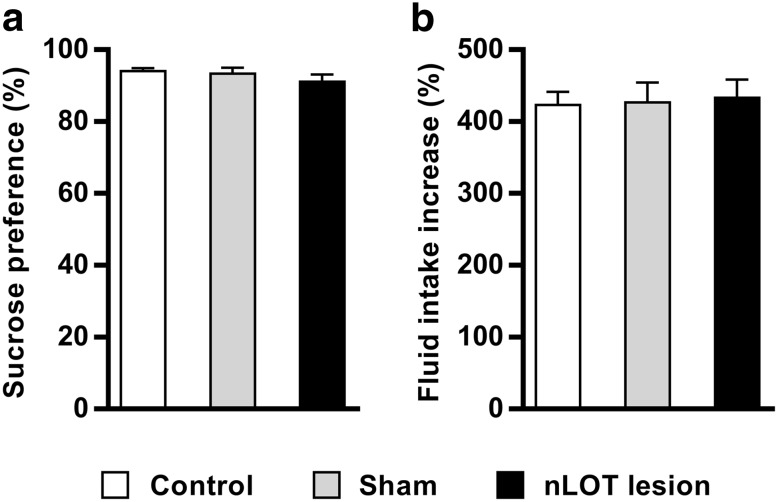
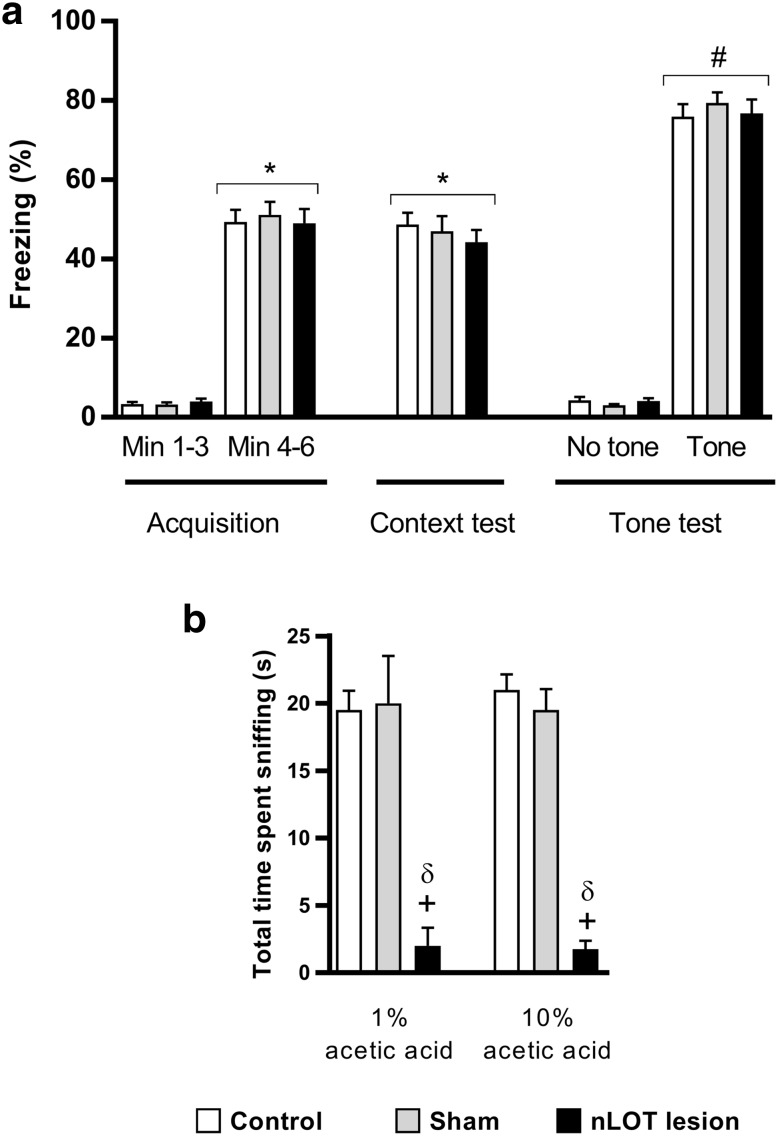
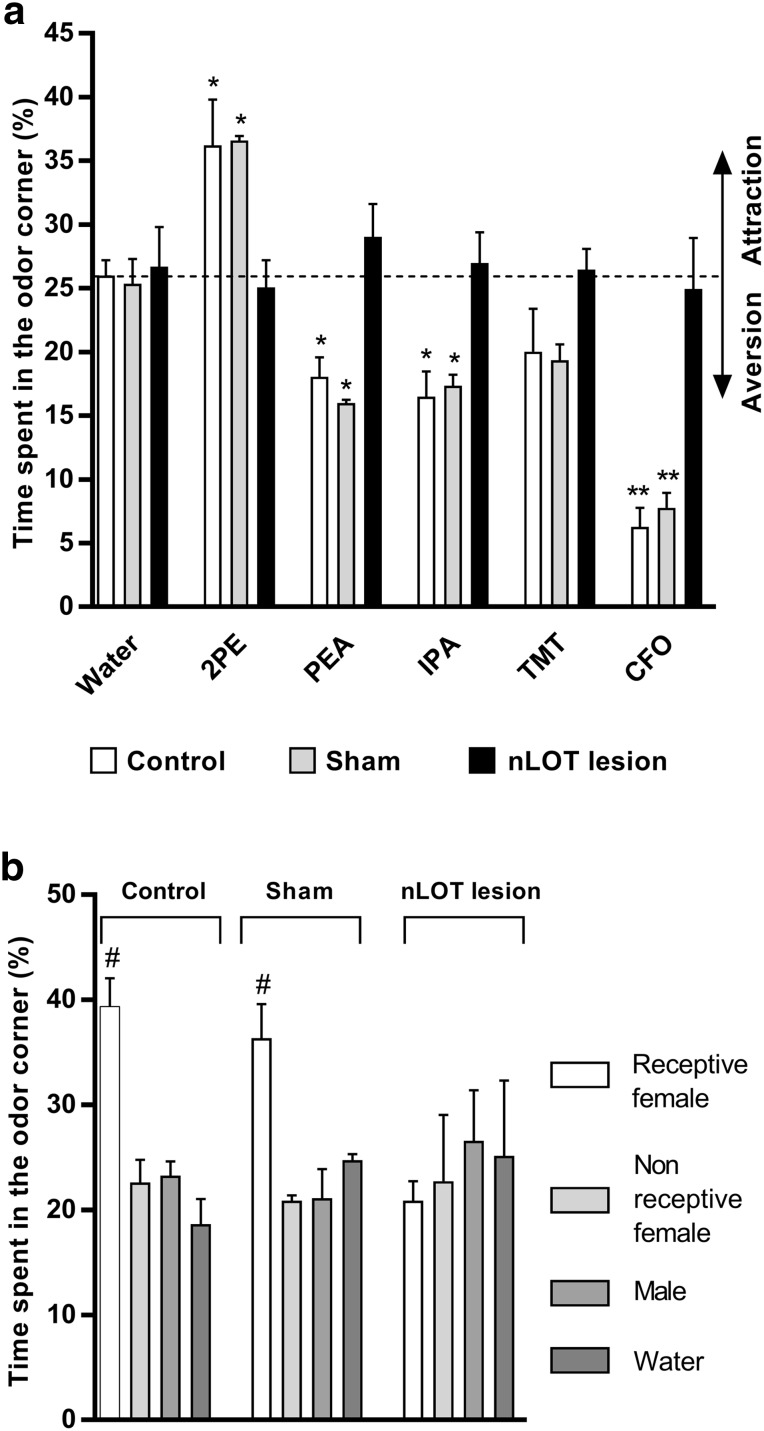
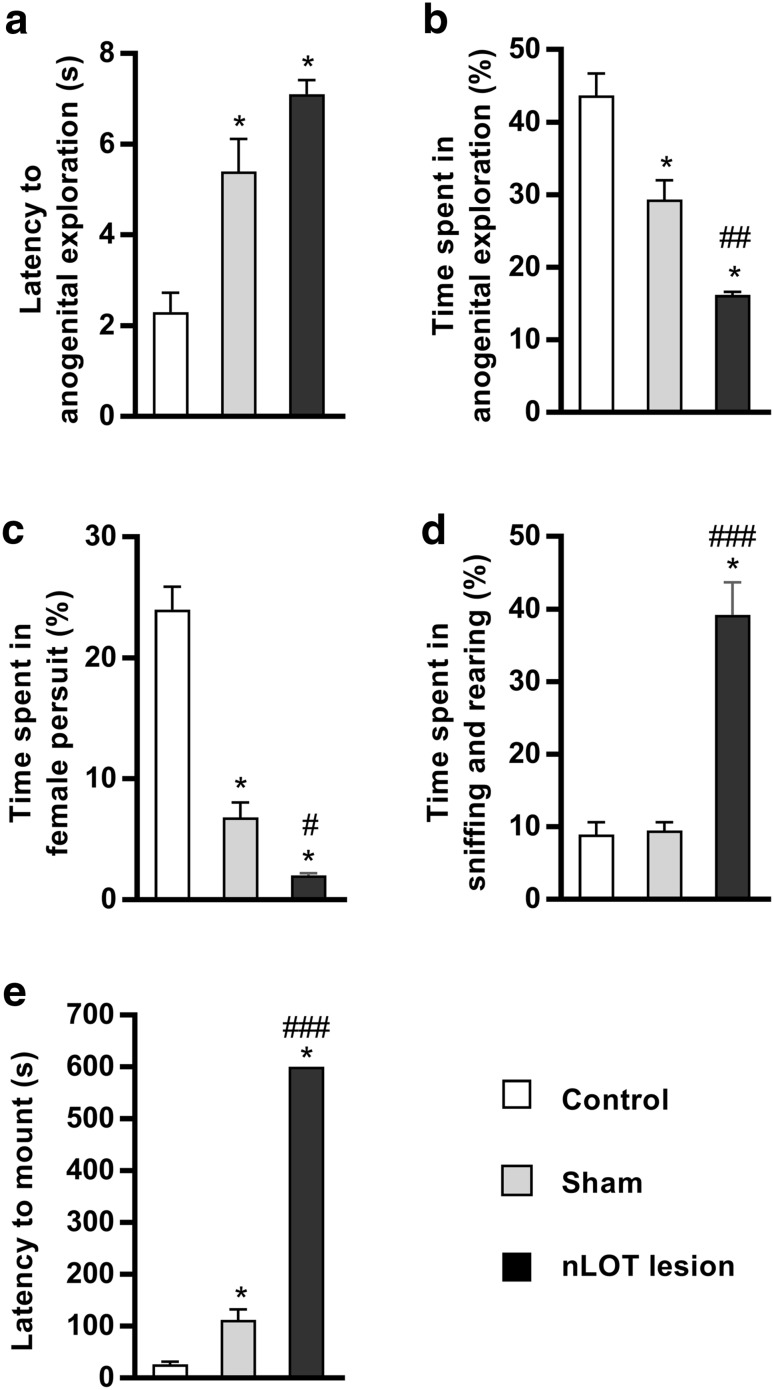
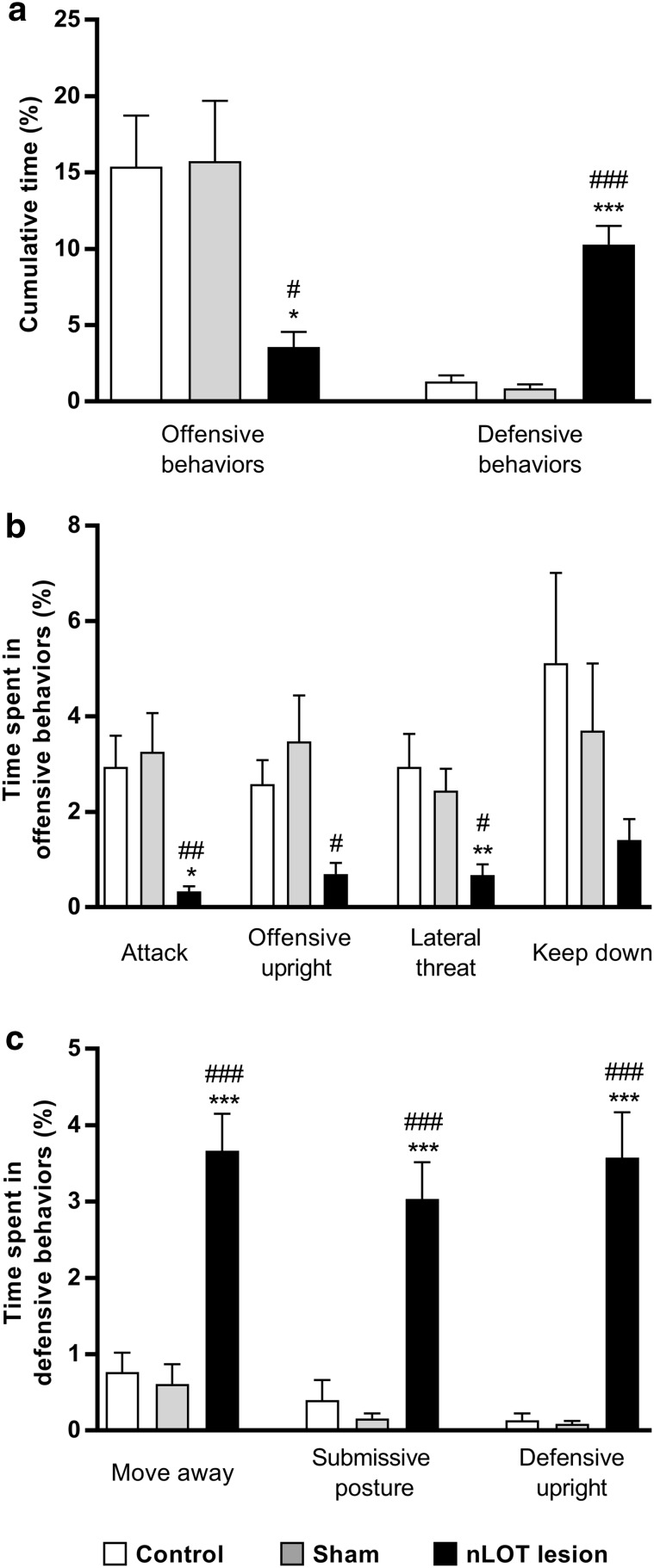
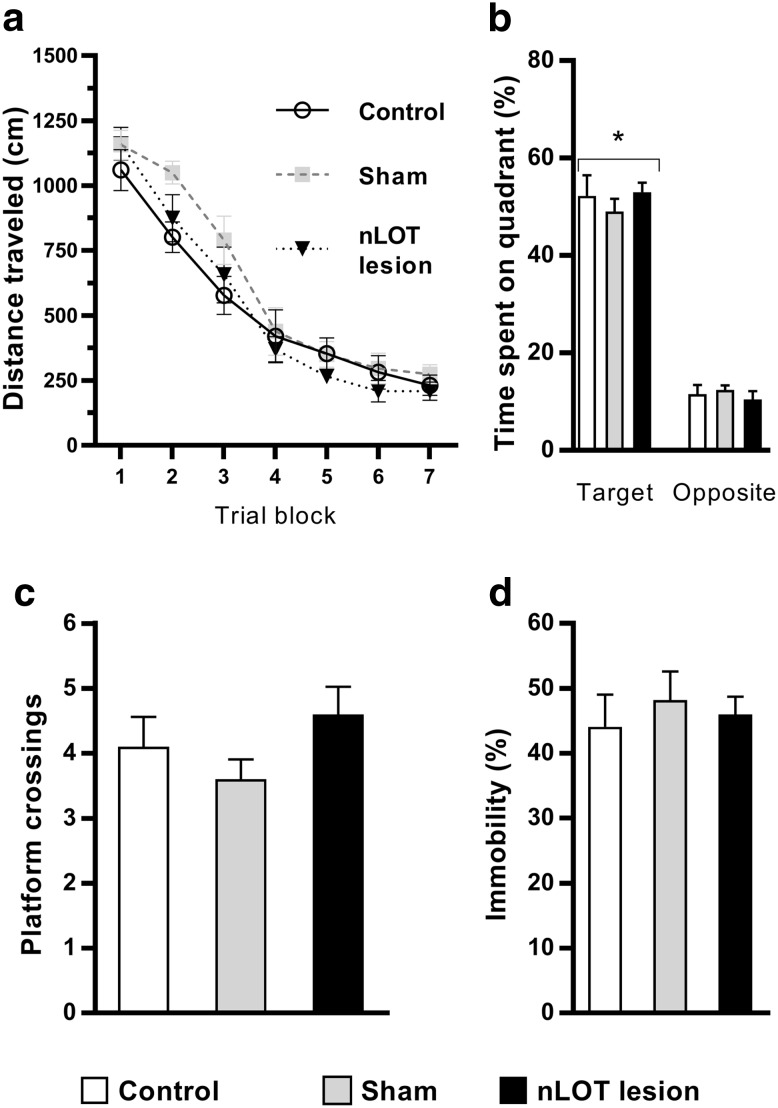
The nucleus of the lateral olfactory tract (nLOT) is a relatively small component of the cortical pallial amygdala, with peculiar neurogenic, neurochemical and connectivity patterns. Although it has been suggested that it might be involved in non-pheromonal olfactory-guided behaviors, particularly feeding, the functional implications of the nLOT have never been investigated. In view of this fact, we have tackled this subject by performing a series of behavioral tests and by quantifying biological and biochemical parameters in sexually naïve adult male rats that were submitted to bilateral excitotoxic lesions of the nLOT. nLOT-lesioned rats had severe olfactory deficits with inability to detect and discriminate between odors. Additionally, they did not display innate behavioral responses to biologically relevant chemosignals. Specifically, nLOT-lesioned rats did not show avoidance towards predator odors or aggressive behaviors towards intruders, and had severely impaired sexual behavior. In fact, nLOT lesions abolished preference for odors of receptive females, reduced chemoinvestigatory behavior and eliminated mounting behavior. nLOT-lesioned rats had normal circulating levels of testosterone, did not display anxiety- or depressive-like behaviors, and had unimpaired cognitive functions and fear acquisition and memory. Altogether, our results suggest that the nLOT integrity is required for the normal functioning of the olfactory system.
Keywords: Aggression; Anosmia; Attractive and avoidance behaviors; Cortical pallial amygdala; Olfactory cortex; Sexual behavior.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
Chronic stress leads to long-lasting deficits in olfactory-guided behaviors, and to neuroplastic changes in the nucleus of the lateral olfactory tract.Horm Behav. 2018 Feb;98:130-144. doi: 10.1016/j.yhbeh.2017.12.006. Epub 2018 Jan 4. Horm Behav. 2018. PMID: 29277699
-
Synaptic Connectivity and Electrophysiological Properties of the Nucleus of the Lateral Olfactory Tract.J Neurosci. 2024 Aug 14;44(33):e2420232024. doi: 10.1523/JNEUROSCI.2420-23.2024. J Neurosci. 2024. PMID: 38997160 Free PMC article.
-
Age effects on the nucleus of the lateral olfactory tract of the rat.J Comp Neurol. 2016 Mar 1;524(4):759-71. doi: 10.1002/cne.23863. Epub 2015 Aug 11. J Comp Neurol. 2016. PMID: 26224652
-
Odor-guided behavior in mammals.Experientia. 1986 Mar 15;42(3):257-71. doi: 10.1007/BF01942506. Experientia. 1986. PMID: 3514263 Review.
-
[Involvement of the olfactory system in learning and memory: a close correlation between the olfactory deficit and the course of Alzheimer's disease?].Yakubutsu Seishin Kodo. 1991 Aug;11(4):223-35. Yakubutsu Seishin Kodo. 1991. PMID: 1799095 Review. Japanese.
Cited by
-
Understanding Emotions: Origins and Roles of the Amygdala.Biomolecules. 2021 May 31;11(6):823. doi: 10.3390/biom11060823. Biomolecules. 2021. PMID: 34072960 Free PMC article. Review.
-
The Zebrafish Amygdaloid Complex - Functional Ground Plan, Molecular Delineation, and Everted Topology.Front Neurosci. 2020 Jul 16;14:608. doi: 10.3389/fnins.2020.00608. eCollection 2020. Front Neurosci. 2020. PMID: 32765204 Free PMC article.
-
Olfactory deficits in aging and Alzheimer's-spotlight on inhibitory interneurons.Front Neurosci. 2024 Dec 16;18:1503069. doi: 10.3389/fnins.2024.1503069. eCollection 2024. Front Neurosci. 2024. PMID: 39737436 Free PMC article. Review.
-
Collateralization patterns of principal basolateral amygdala neurons delineate distinct output streams.iScience. 2025 Jul 11;28(8):113089. doi: 10.1016/j.isci.2025.113089. eCollection 2025 Aug 15. iScience. 2025. PMID: 40799396 Free PMC article.
-
A whole-brain atlas of monosynaptic input targeting four different cell types in the medial prefrontal cortex of the mouse.Nat Neurosci. 2019 Apr;22(4):657-668. doi: 10.1038/s41593-019-0354-y. Epub 2019 Mar 18. Nat Neurosci. 2019. PMID: 30886408
References
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
