

iPSC-Derived Human Microglia-like Cells to Study Neurological Diseases
- PMID: 28426964
- PMCID: PMC5482419
- DOI: 10.1016/j.neuron.2017.03.042
iPSC-Derived Human Microglia-like Cells to Study Neurological Diseases
Abstract
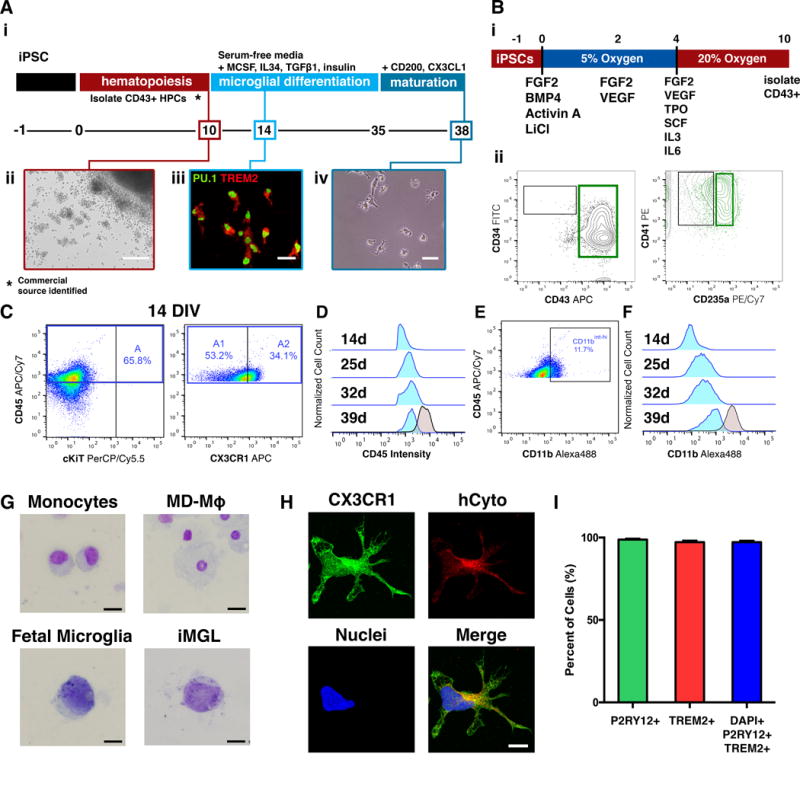
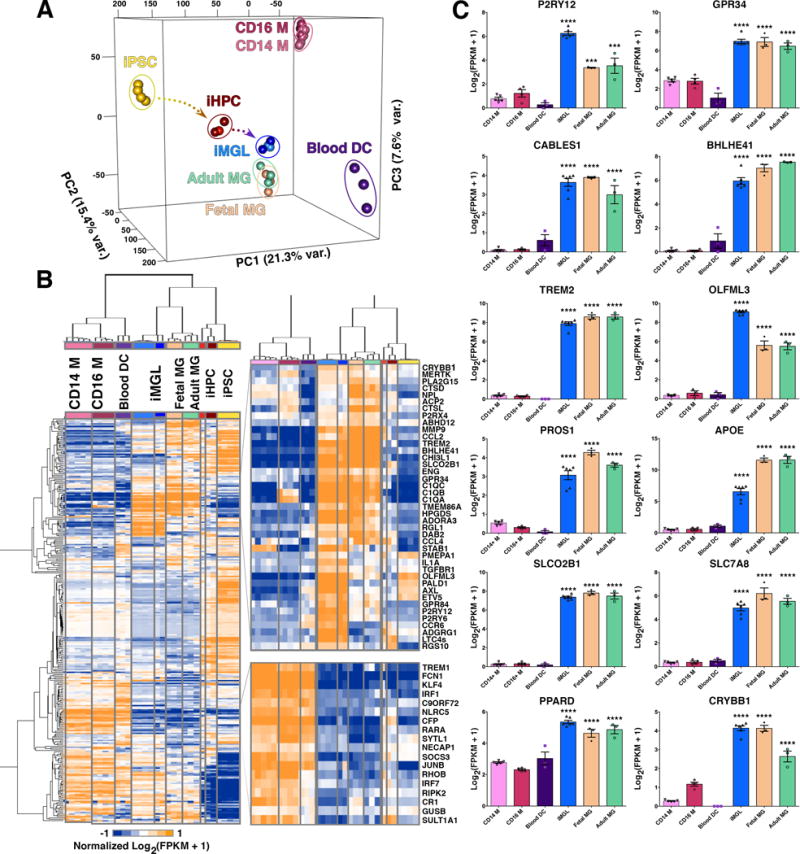
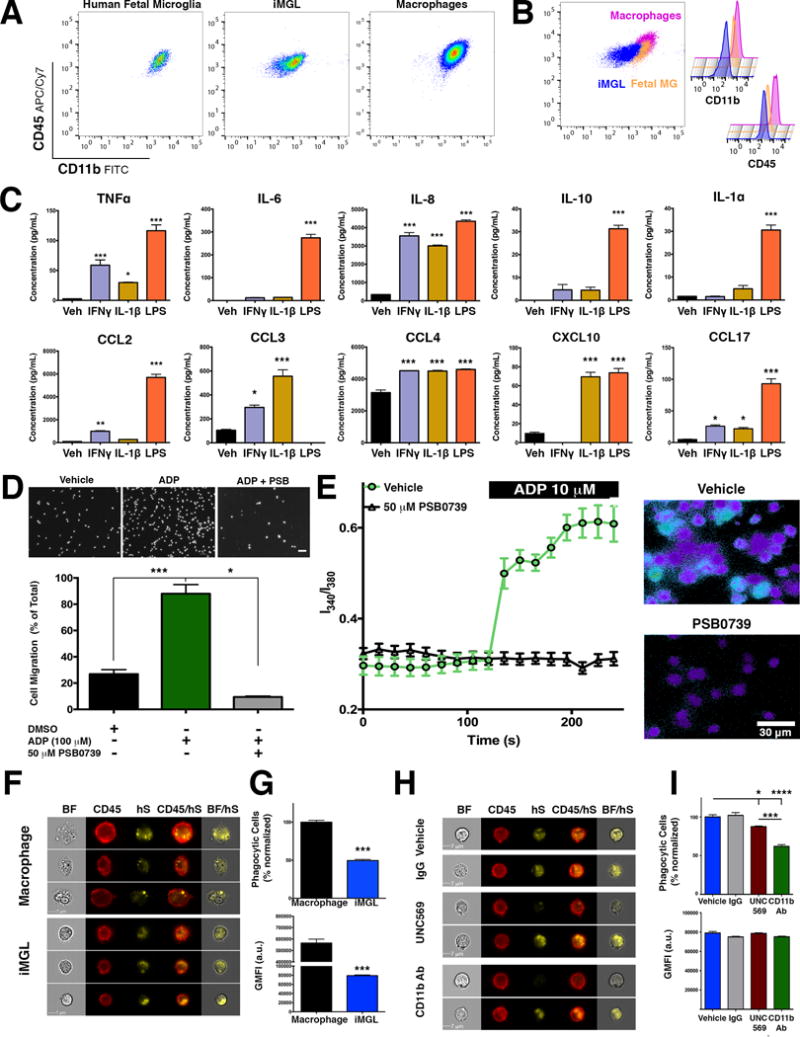
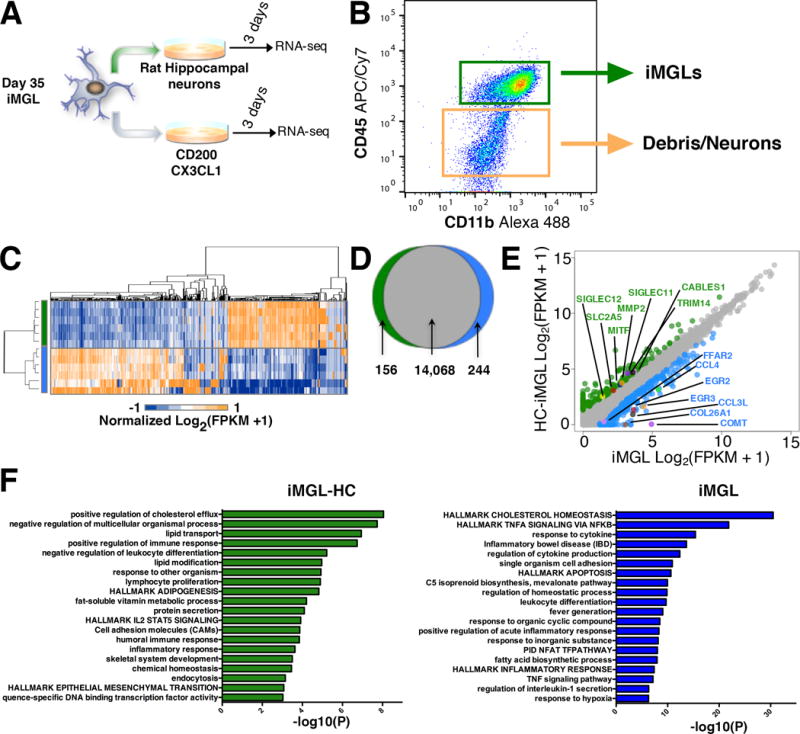
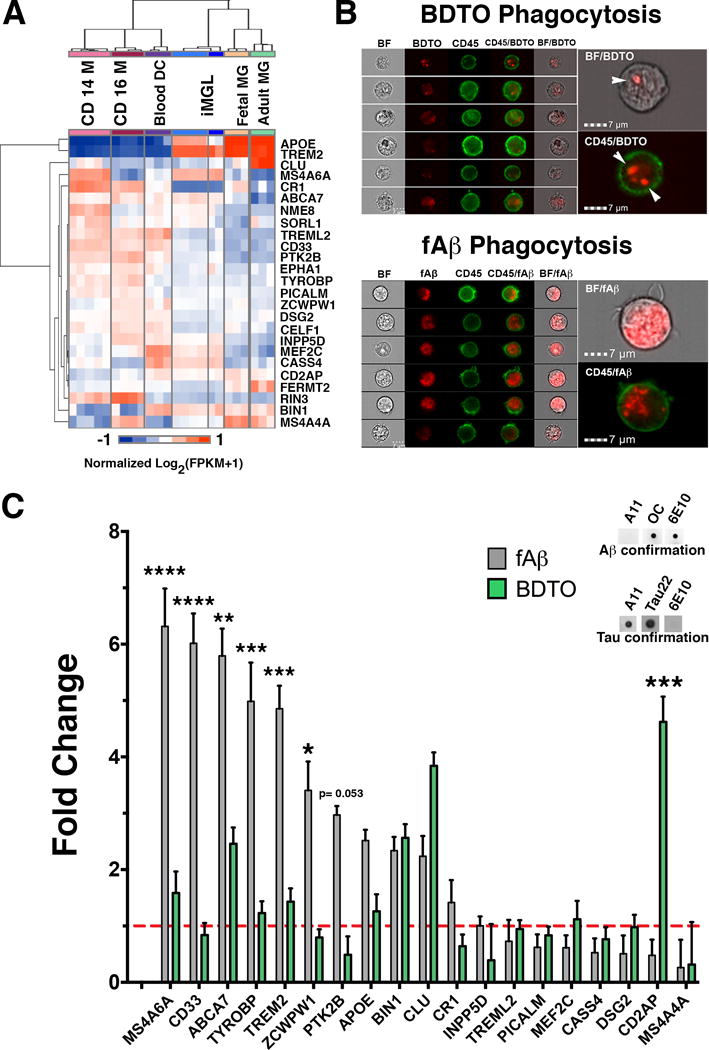
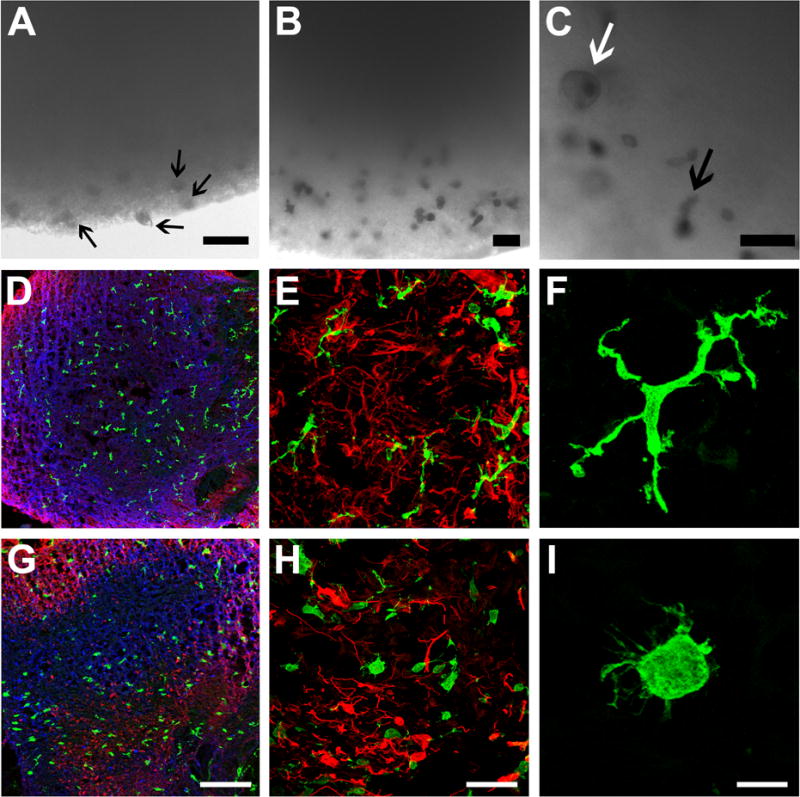
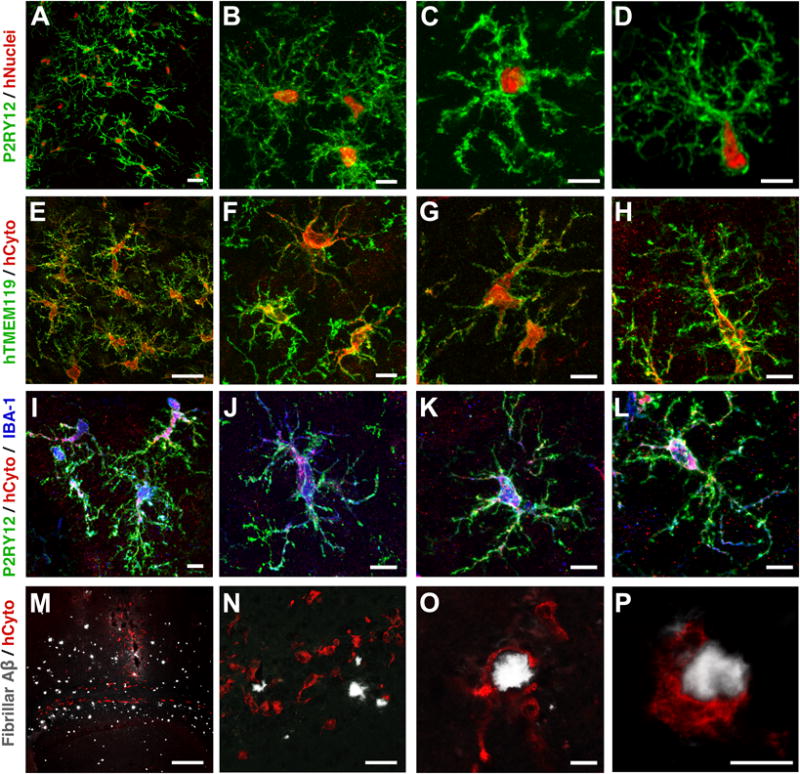
Microglia play critical roles in brain development, homeostasis, and neurological disorders. Here, we report that human microglial-like cells (iMGLs) can be differentiated from iPSCs to study their function in neurological diseases, like Alzheimer's disease (AD). We find that iMGLs develop in vitro similarly to microglia in vivo, and whole-transcriptome analysis demonstrates that they are highly similar to cultured adult and fetal human microglia. Functional assessment of iMGLs reveals that they secrete cytokines in response to inflammatory stimuli, migrate and undergo calcium transients, and robustly phagocytose CNS substrates. iMGLs were used to examine the effects of Aβ fibrils and brain-derived tau oligomers on AD-related gene expression and to interrogate mechanisms involved in synaptic pruning. Furthermore, iMGLs transplanted into transgenic mice and human brain organoids resemble microglia in vivo. Together, these findings demonstrate that iMGLs can be used to study microglial function, providing important new insight into human neurological disease.
Keywords: 3D organoids; AD-GWAS; Alzheimer’s disease; Beta-amyloid; Tau; cell models of disease; induced pluripotent stem cells; microglia; mouse transplantation; neurodegenerative diseases.
Copyright © 2017 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no conflict of interests.
Figures
References
-
- Abbas N, Bednar I, Mix E, Marie S, Paterson D, Ljungberg A, Morris C, Winblad B, Nordberg A, Zhu J. Up-regulation of the inflammatory cytokines IFN-gamma and IL-12 and down-regulation of IL-4 in cerebral cortex regions of APP(SWE) transgenic mice. J Neuroimmunol. 2002;126:50–57. - PubMed
-
- Abdollahi A, Lord KA, Hoffman-Liebermann B, Liebermann DA. Interferon regulatory factor 1 is a myeloid differentiation primary response gene induced by interleukin 6 and leukemia inhibitory factor: role in growth inhibition. Cell Growth Differ. 1991;2:401–407. - PubMed
-
- Abutbul S, Shapiro J, Szaingurten-Solodkin I, Levy N, Carmy Y, Baron R, Jung S, Monsonego A. TGF-beta signaling through SMAD2/3 induces the quiescent microglial phenotype within the CNS environment. Glia. 2012;60:1160–1171. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
