

Assembly of Excitatory Synapses in the Absence of Glutamatergic Neurotransmission
- PMID: 28426966
- PMCID: PMC5521186
- DOI: 10.1016/j.neuron.2017.03.047
Assembly of Excitatory Synapses in the Absence of Glutamatergic Neurotransmission
Abstract
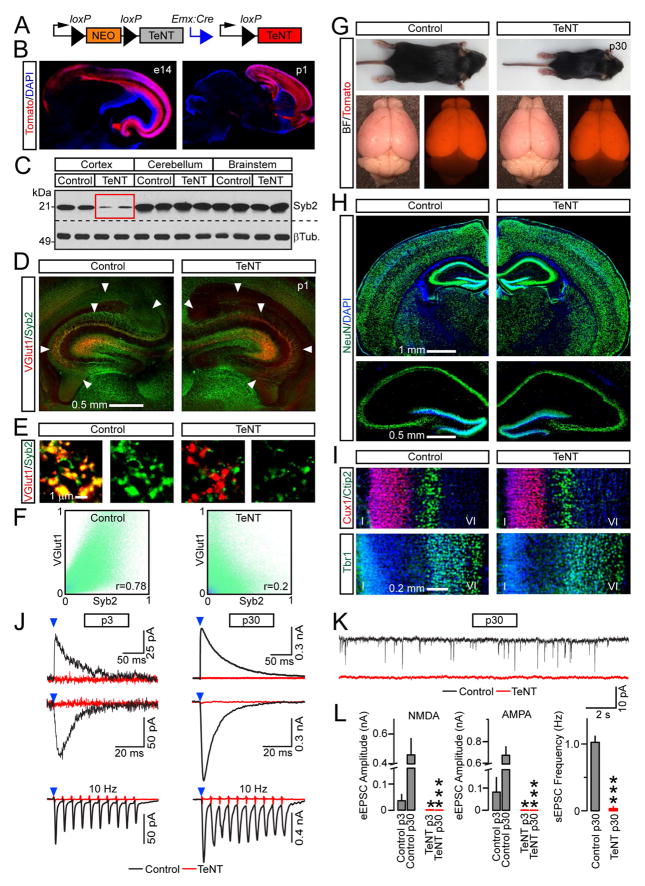
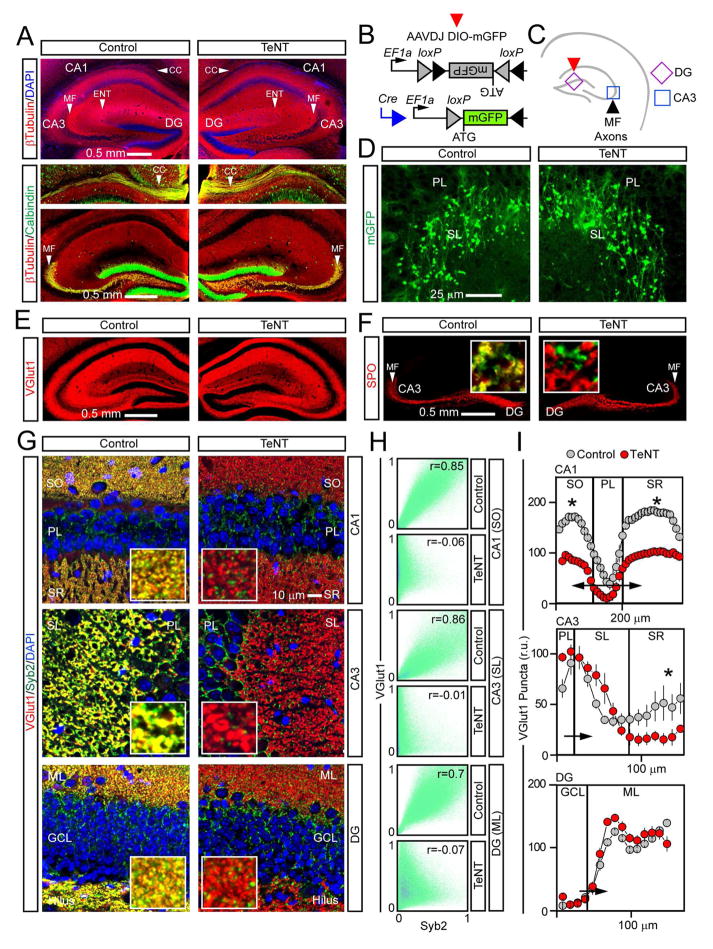
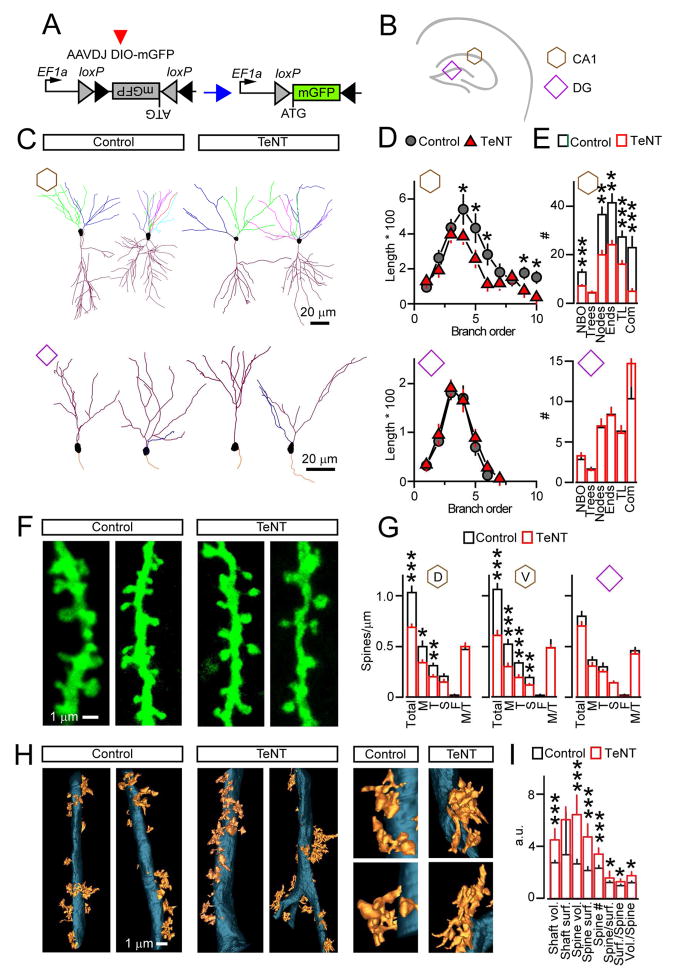
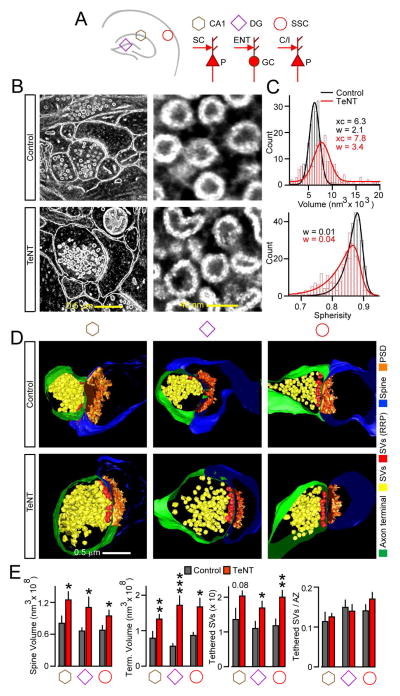
Synaptic excitation mediates a broad spectrum of structural changes in neural circuits across the brain. Here, we examine the morphologies, wiring, and architectures of single synapses of projection neurons in the murine hippocampus that developed in virtually complete absence of vesicular glutamate release. While these neurons had smaller dendritic trees and/or formed fewer contacts in specific hippocampal subfields, their stereotyped connectivity was largely preserved. Furthermore, loss of release did not disrupt the morphogenesis of presynaptic terminals and dendritic spines, suggesting that glutamatergic neurotransmission is unnecessary for synapse assembly and maintenance. These results underscore the instructive role of intrinsic mechanisms in synapse formation.
Keywords: active zone; dendritic spine; glutamate release; hardwiring; hippocampus; memory; neural circuit development; neurotransmission; synapse; synaptic vesicle exocytosis.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
References
-
- Alvarez VA, Sabatini BL. Anatomical and physiological plasticity of dendritic spines. Annual review of neuroscience. 2007;30:79–97. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
