

Modulation of Asymmetric Flux in Heterotypic Gap Junctions by Pore Shape, Particle Size and Charge
- PMID: 28428758
- PMCID: PMC5382223
- DOI: 10.3389/fphys.2017.00206
Modulation of Asymmetric Flux in Heterotypic Gap Junctions by Pore Shape, Particle Size and Charge
Abstract
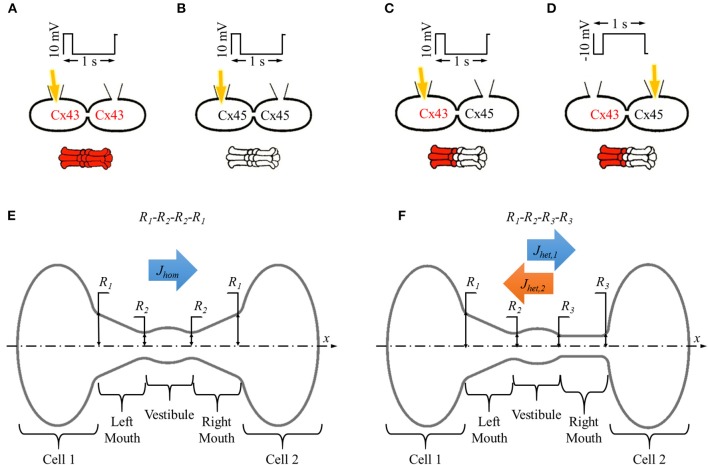
Gap junction channels play a vital role in intercellular communication by connecting cytoplasm of adjoined cells through arrays of channel-pores formed at the common membrane junction. Their structure and properties vary depending on the connexin isoform(s) involved in forming the full gap junction channel. Lack of information on the molecular structure of gap junction channels has limited the development of computational tools for single channel studies. Currently, we rely on cumbersome experimental techniques that have limited capabilities. We have earlier reported a simplified Brownian dynamics gap junction pore model and demonstrated that variations in pore shape at the single channel level can explain some of the differences in permeability of heterotypic channels observed in in vitro experiments. Based on this computational model, we designed simulations to study the influence of pore shape, particle size and charge in homotypic and heterotypic pores. We simulated dye diffusion under whole cell voltage clamping. Our simulation studies with pore shape variations revealed a pore shape with maximal flux asymmetry in a heterotypic pore. We identified pore shape profiles that match the in silico flux asymmetry results to the in vitro results of homotypic and heterotypic gap junction formed out of Cx43 and Cx45. Our simulation results indicate that the channel's pore-shape established flux asymmetry and that flux asymmetry is primarily regulated by the sizes of the conical and/or cylindrical mouths at each end of the pore. Within the set range of particle size and charge, flux asymmetry was found to be independent of particle size and directly proportional to charge magnitude. While particle charge was vital to creating flux asymmetry, charge magnitude only scaled the observed flux asymmetry. Our studies identified the key factors that help predict asymmetry. Finally, we suggest the role of such flux asymmetry in creating concentration imbalances of messenger molecules in cardiomyocytes. We also assess the potency of fibroblasts in aggravating such imbalances through Cx43-Cx45 heterotypic channels in fibrotic heart tissue.
Keywords: Brownian dynamics; diffusion simulation; heterotypic gap junctions; intercellular communication; mathematical modeling; permeability.
Figures






Similar articles
-
Computational simulations of asymmetric fluxes of large molecules through gap junction channel pores.J Theor Biol. 2017 Jan 7;412:61-73. doi: 10.1016/j.jtbi.2016.08.040. Epub 2016 Aug 31. J Theor Biol. 2017. PMID: 27590324
-
The permeability of gap junction channels to probes of different size is dependent on connexin composition and permeant-pore affinities.Biophys J. 2004 Aug;87(2):958-73. doi: 10.1529/biophysj.103.036350. Biophys J. 2004. PMID: 15298902 Free PMC article.
-
Functional formation of heterotypic gap junction channels by connexins-40 and -43.Channels (Austin). 2014;8(5):433-43. doi: 10.4161/19336950.2014.949188. Channels (Austin). 2014. PMID: 25483586 Free PMC article.
-
Emerging issues of connexin channels: biophysics fills the gap.Q Rev Biophys. 2001 Aug;34(3):325-472. doi: 10.1017/s0033583501003705. Q Rev Biophys. 2001. PMID: 11838236 Review.
-
Size and selectivity of gap junction channels formed from different connexins.J Bioenerg Biomembr. 1996 Aug;28(4):327-37. doi: 10.1007/BF02110109. J Bioenerg Biomembr. 1996. PMID: 8844330 Review.
Cited by
-
Connexins and Pannexins: Important Players in Neurodevelopment, Neurological Diseases, and Potential Therapeutics.Biomedicines. 2022 Sep 9;10(9):2237. doi: 10.3390/biomedicines10092237. Biomedicines. 2022. PMID: 36140338 Free PMC article. Review.
References
-
- Allen T. W., Hoyles M., Kuyucak S., Chung S. H. (1999). Molecular and Brownian dynamics study of ion selectivity and conductivity in the potassium channel. Chem. Phys. Lett. 313, 358–365. 10.1016/S0009-2614(99)01004-0 - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous