

Quantifying Golgi structure using EM: combining volume-SEM and stereology for higher throughput
- PMID: 28429122
- PMCID: PMC5429891
- DOI: 10.1007/s00418-017-1564-6
Quantifying Golgi structure using EM: combining volume-SEM and stereology for higher throughput
Abstract
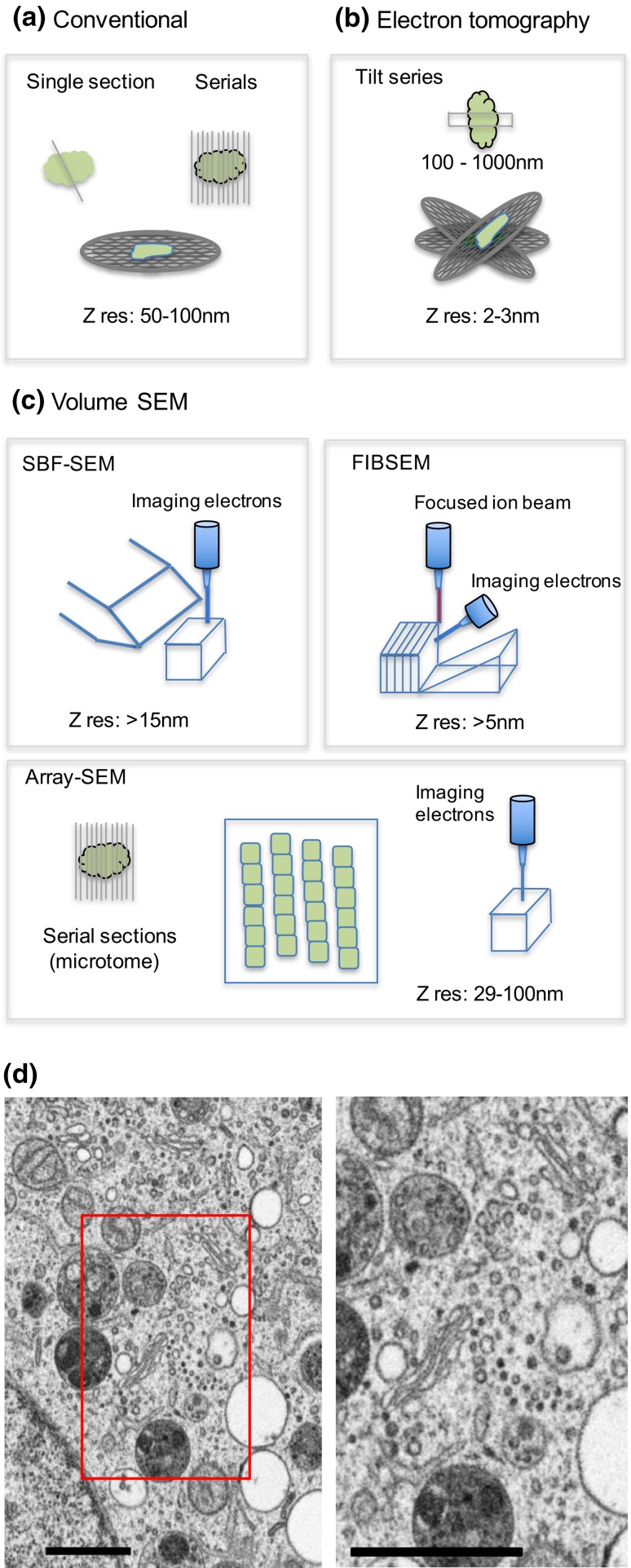
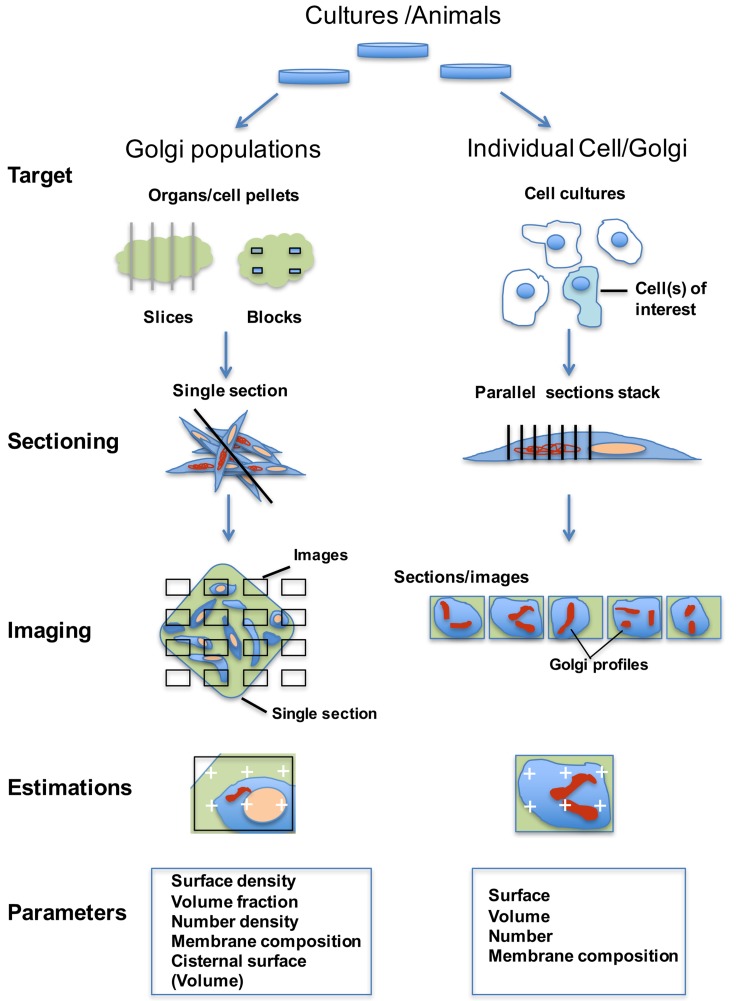
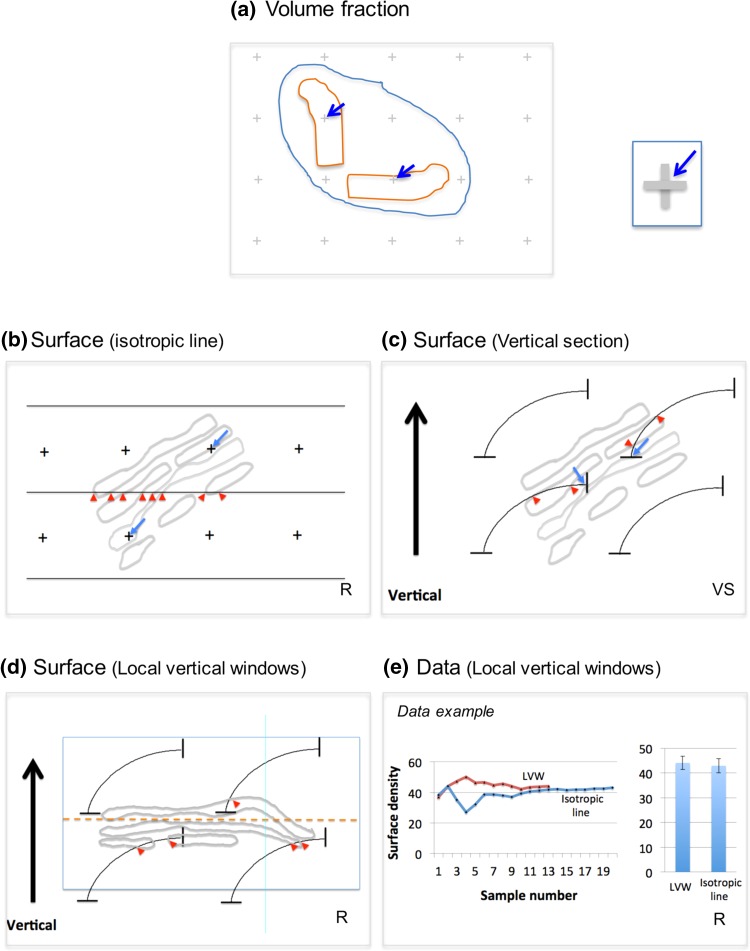
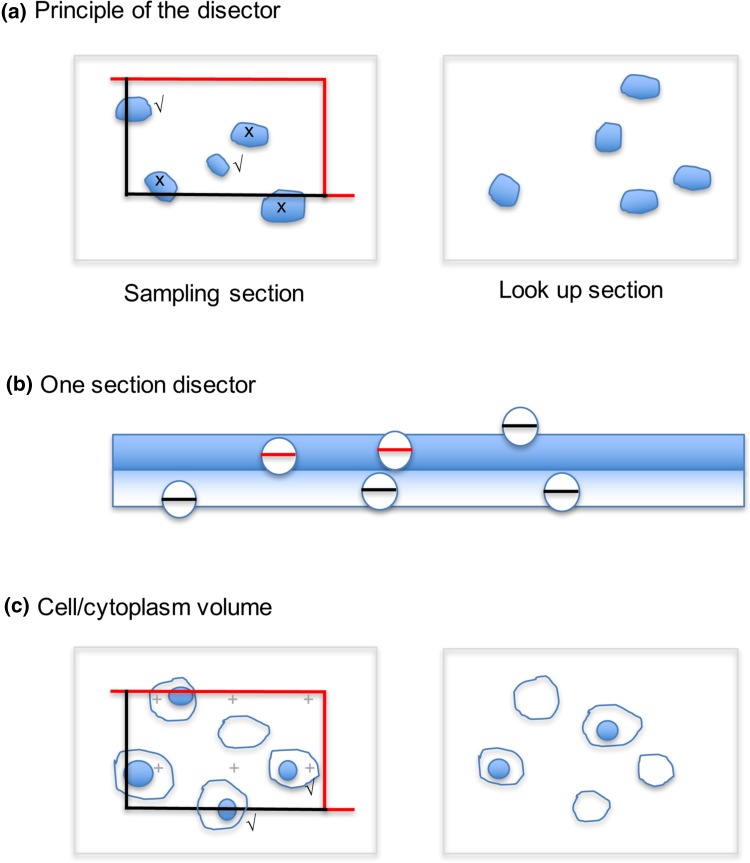
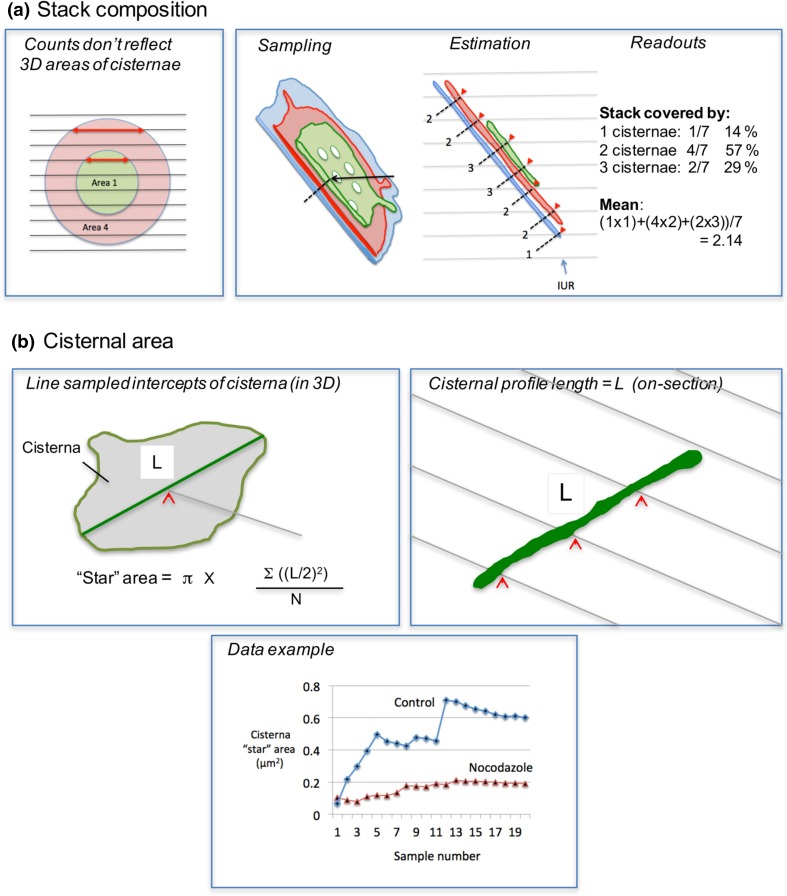
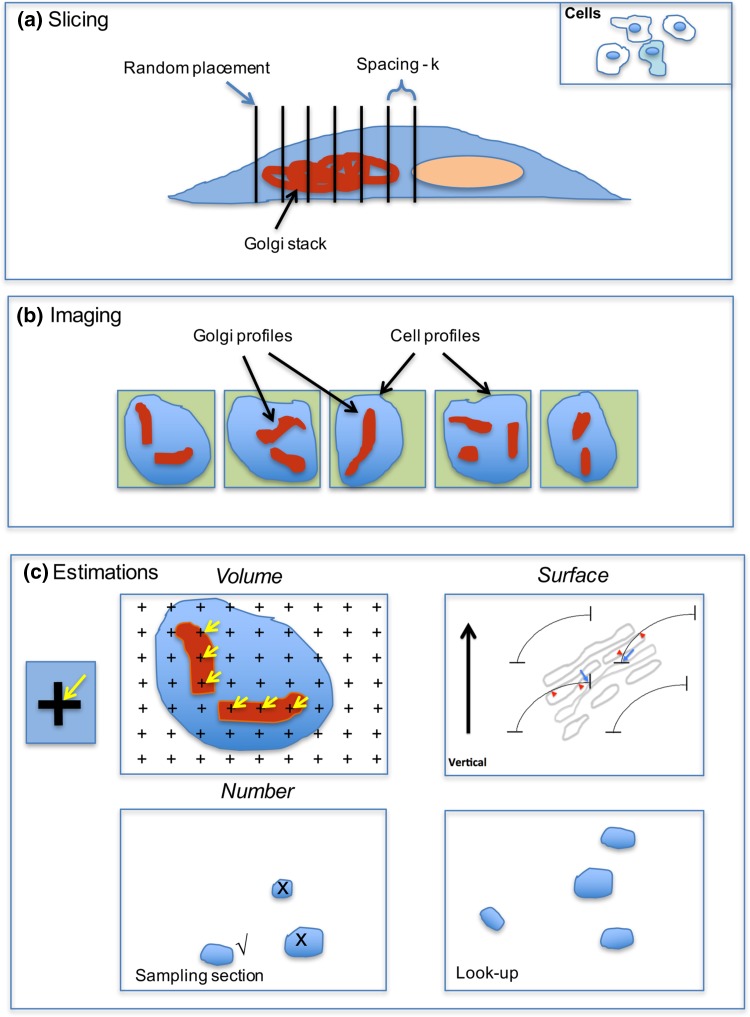
Investigating organelles such as the Golgi complex depends increasingly on high-throughput quantitative morphological analyses from multiple experimental or genetic conditions. Light microscopy (LM) has been an effective tool for screening but fails to reveal fine details of Golgi structures such as vesicles, tubules and cisternae. Electron microscopy (EM) has sufficient resolution but traditional transmission EM (TEM) methods are slow and inefficient. Newer volume scanning EM (volume-SEM) methods now have the potential to speed up 3D analysis by automated sectioning and imaging. However, they produce large arrays of sections and/or images, which require labour-intensive 3D reconstruction for quantitation on limited cell numbers. Here, we show that the information storage, digital waste and workload involved in using volume-SEM can be reduced substantially using sampling-based stereology. Using the Golgi as an example, we describe how Golgi populations can be sensed quantitatively using single random slices and how accurate quantitative structural data on Golgi organelles of individual cells can be obtained using only 5-10 sections/images taken from a volume-SEM series (thereby sensing population parameters and cell-cell variability). The approach will be useful in techniques such as correlative LM and EM (CLEM) where small samples of cells are treated and where there may be variable responses. For Golgi study, we outline a series of stereological estimators that are suited to these analyses and suggest workflows, which have the potential to enhance the speed and relevance of data acquisition in volume-SEM.
Keywords: FIBSEM; Golgi; Quantification; SBF-SEM; Sampling; Stereology; Volume-SEM.
Figures

References
-
- Baddeley A, Jensen EBV. Stereology for statisticians. Boca Raton: Chapman and Hall/CRC press; 2004. p. 23.
-
- Ballerini M, Milani M, Costato M, Squadrini F, Turcu IC. Life science applications of focused ion beams (FIB) Eur J Histochem. 1997;41(Suppl 2):89–90. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
