

Parkin regulates translesion DNA synthesis in response to UV radiation
- PMID: 28430587
- PMCID: PMC5482665
- DOI: 10.18632/oncotarget.16855
Parkin regulates translesion DNA synthesis in response to UV radiation
Abstract
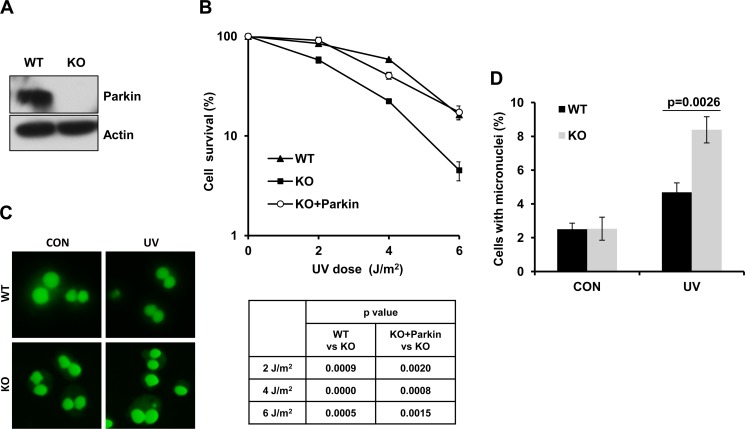
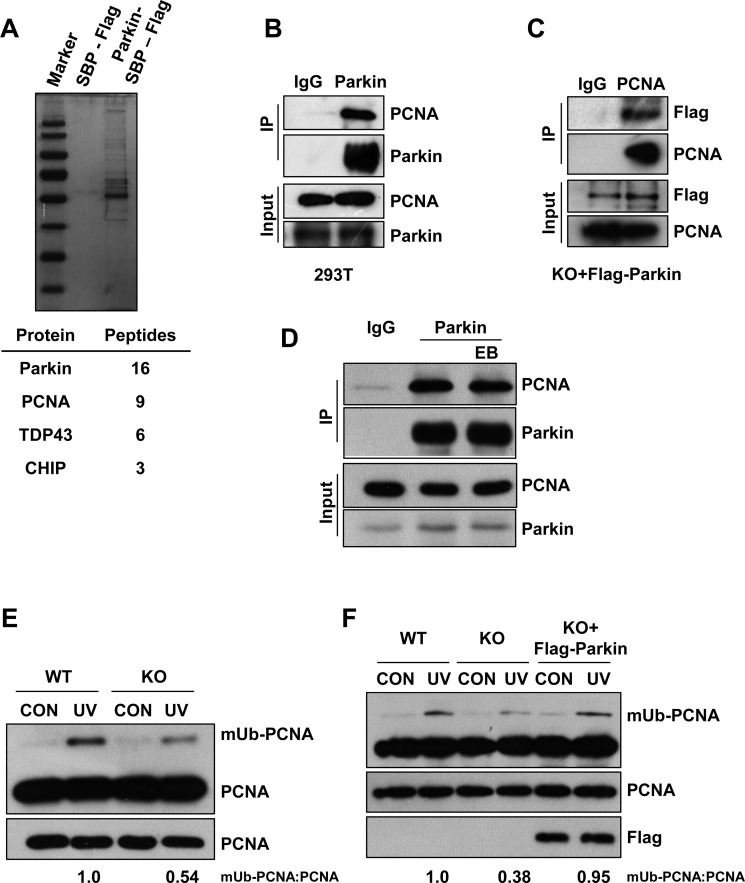
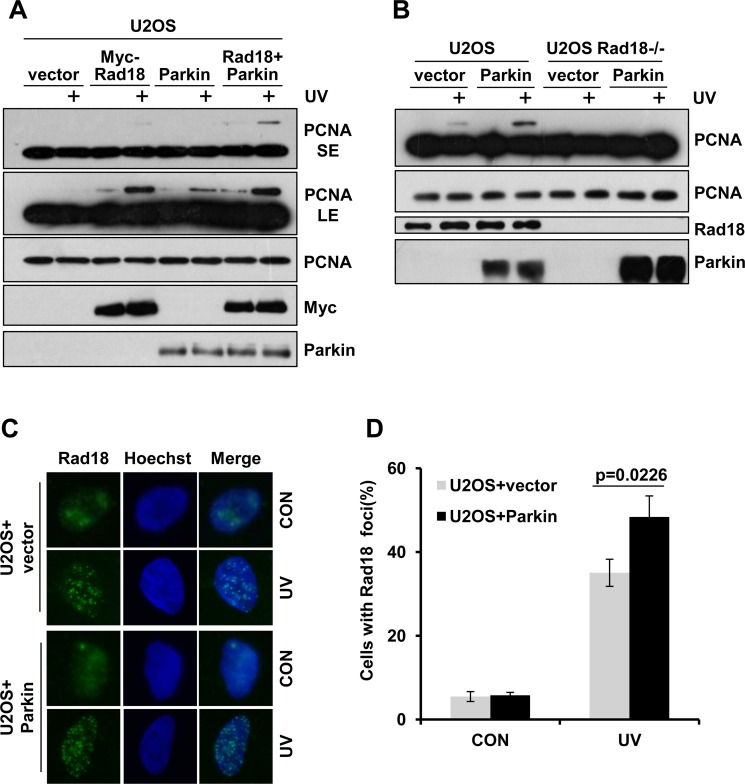
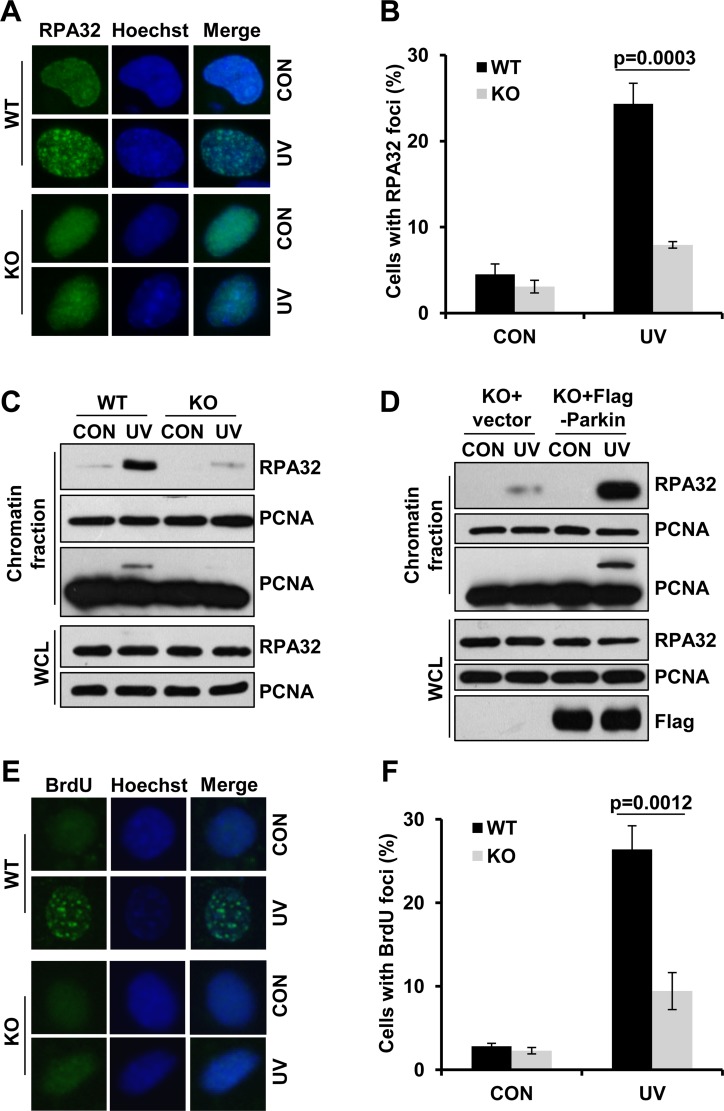
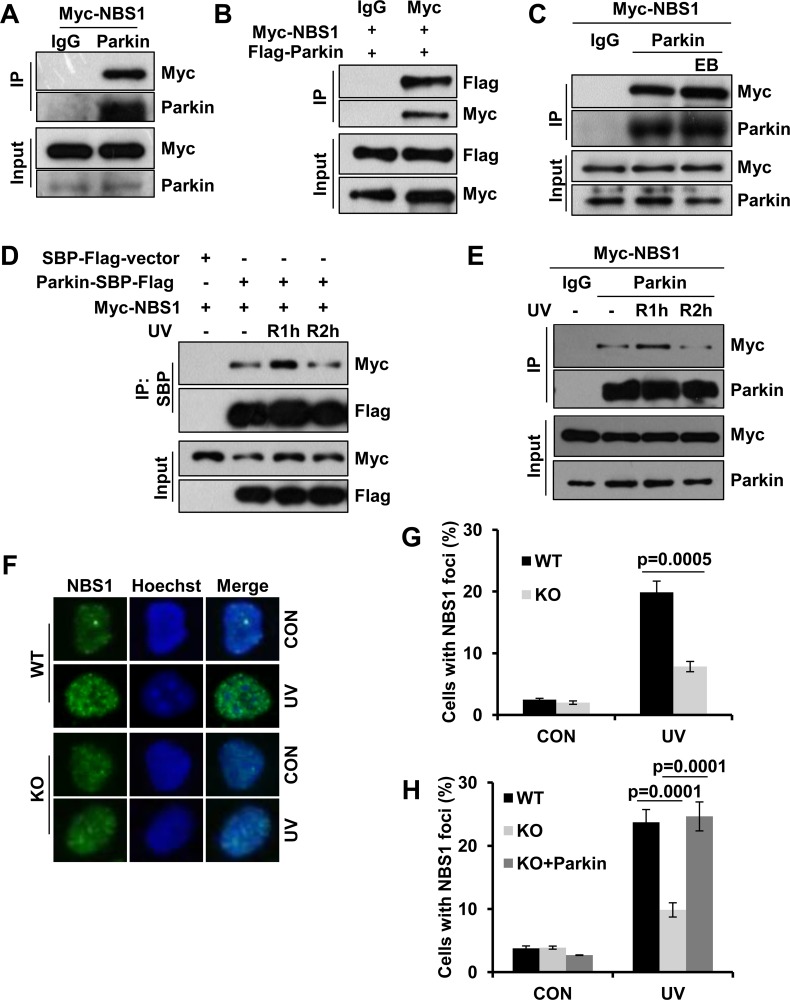
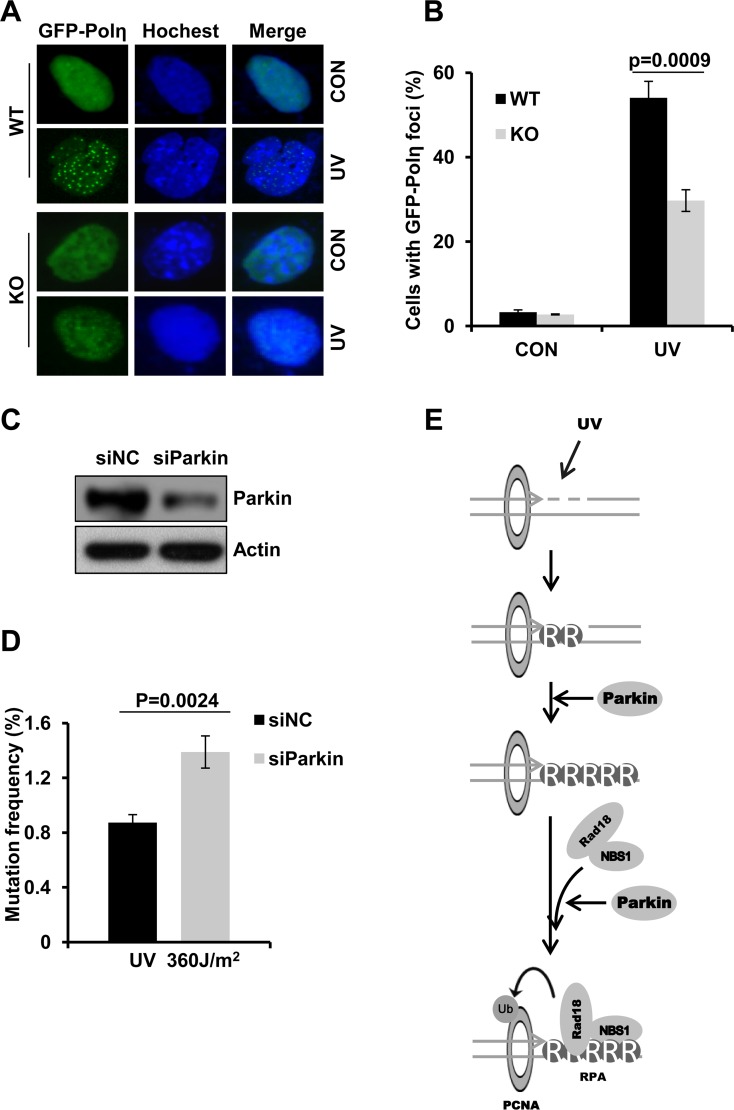
Deficiency of Parkin is a major cause of early-onset Parkinson's disease (PD). Notably, PD patients also exhibit a significantly higher risk in melanoma and other skin tumors, while the mechanism remains largely unknown. In this study, we show that depletion of Parkin causes compromised cell viability and genome stability after ultraviolet (UV) radiation. We demonstrate that Parkin promotes efficient Rad18-dependent proliferating cell nuclear antigen (PCNA) monoubiquitination by facilitating the formation of Replication protein A (RPA)-coated ssDNA upon UV radiation. Furthermore, Parkin is found to physically interact with NBS1 (Nijmegen breakage syndrome 1), and to be required for optimal recruitment of NBS1 and DNA polymerase eta (Polη) to UV-induced damage sites. Consequently, depletion of Parkin leads to increased UV-induced mutagenesis. These findings unveil an important role of Parkin in protecting genome stability through positively regulating translesion DNA synthesis (TLS) upon UV damage, providing a novel mechanistic link between Parkin deficiency and predisposition to skin cancers in PD patients.
Keywords: Parkin; Parkinson’s disease; melanoma; translesion DNA synthesis; ultraviolet radiation.
Conflict of interest statement
The authors declare no conflicts of interests.
Figures
Similar articles
-
RNA-splicing factor SART3 regulates translesion DNA synthesis.Nucleic Acids Res. 2018 May 18;46(9):4560-4574. doi: 10.1093/nar/gky220. Nucleic Acids Res. 2018. PMID: 29590477 Free PMC article.
-
NBS1 recruits RAD18 via a RAD6-like domain and regulates Pol η-dependent translesion DNA synthesis.Mol Cell. 2011 Sep 2;43(5):788-97. doi: 10.1016/j.molcel.2011.07.026. Mol Cell. 2011. PMID: 21884979
-
Replication protein A dynamically regulates monoubiquitination of proliferating cell nuclear antigen.J Biol Chem. 2019 Mar 29;294(13):5157-5168. doi: 10.1074/jbc.RA118.005297. Epub 2019 Jan 30. J Biol Chem. 2019. PMID: 30700555 Free PMC article.
-
Eukaryotic DNA damage tolerance and translesion synthesis through covalent modifications of PCNA.Cell Res. 2008 Jan;18(1):162-73. doi: 10.1038/cr.2007.114. Cell Res. 2008. PMID: 18157158 Review.
-
CRL4Cdt2: master coordinator of cell cycle progression and genome stability.Cell Cycle. 2011 Jan 15;10(2):241-9. doi: 10.4161/cc.10.2.14530. Epub 2011 Jan 15. Cell Cycle. 2011. PMID: 21212733 Free PMC article. Review.
Cited by
-
More than Meets the ISG15: Emerging Roles in the DNA Damage Response and Beyond.Biomolecules. 2020 Nov 15;10(11):1557. doi: 10.3390/biom10111557. Biomolecules. 2020. PMID: 33203188 Free PMC article. Review.
-
Decoding the Role of Familial Parkinson's Disease-Related Genes in DNA Damage and Repair.Aging Dis. 2022 Oct 1;13(5):1405-1412. doi: 10.14336/AD.2022.0216. eCollection 2022 Oct 1. Aging Dis. 2022. PMID: 36186134 Free PMC article.
-
RNA-splicing factor SART3 regulates translesion DNA synthesis.Nucleic Acids Res. 2018 May 18;46(9):4560-4574. doi: 10.1093/nar/gky220. Nucleic Acids Res. 2018. PMID: 29590477 Free PMC article.
-
Link between Parkinson's disease and melanoma: insights into the influence of the PARK gene family.Front Oncol. 2025 Aug 11;15:1506744. doi: 10.3389/fonc.2025.1506744. eCollection 2025. Front Oncol. 2025. PMID: 40860809 Free PMC article. Review.
References
-
- Lucking CB, Durr A, Bonifati V, Vaughan J, De Michele G, Gasser T, Harhangi BS, Meco G, Denefle P, Wood NW, Agid Y, Brice A, French Parkinson's Disease Genetics Study G et al. Association between early-onset Parkinson's disease and mutations in the parkin gene. N Engl J Med. 2000;342:1560–7. doi: 10.1056/NEJM200005253422103. - DOI - PubMed
-
- Riley BE, Lougheed JC, Callaway K, Velasquez M, Brecht E, Nguyen L, Shaler T, Walker D, Yang Y, Regnstrom K, Diep L, Zhang Z, Chiou S, et al. Structure and function of Parkin E3 ubiquitin ligase reveals aspects of RING and HECT ligases. Nat Commun. 2013;4:1982. doi: 10.1038/ncomms2982. - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
