

Role of phosphatase and tensin homolog in hypoxic pulmonary vasoconstriction
- PMID: 28430879
- PMCID: PMC5852630
- DOI: 10.1093/cvr/cvx076
Role of phosphatase and tensin homolog in hypoxic pulmonary vasoconstriction
Abstract
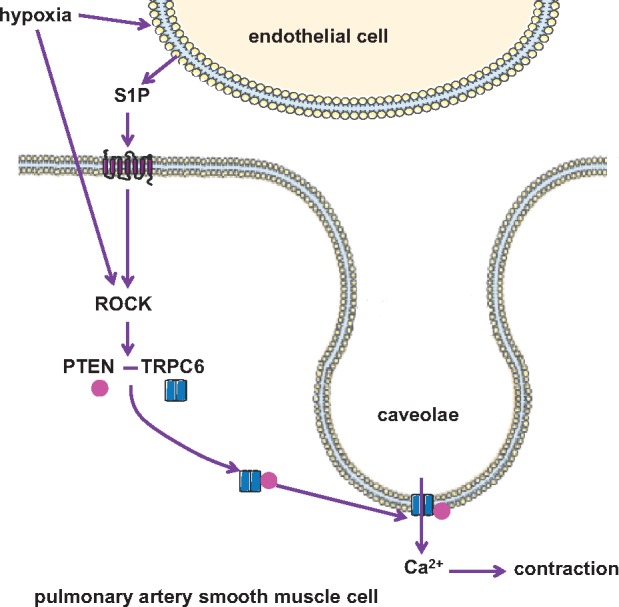
Aims: Hypoxic pulmonary vasoconstriction (HPV) redistributes blood flow from poorly ventilated to better aerated areas in the lung, thereby optimizing ventilation-perfusion ratio (V/Q). Pulmonary artery smooth muscle cell (PASMC) contraction in response to hypoxia is triggered by Ca2+ influx via transient receptor potential canonical 6 (TRPC6) cation channels that have translocated to caveolae in the plasma membrane. Since phosphatase and tensin homolog (PTEN) was suggested to regulate TRPC6 in endothelial cells, we aimed to define its role in the hypoxic response of PASMCs and as a putative mediator of HPV.
Methods and results: In isolated perfused mouse lungs, smooth muscle specific PTEN deficiency attenuated pulmonary vasoconstriction in response to hypoxia but not to angiotensin II (Ang II). Analogously, siRNA-mediated knock down of PTEN in human PASMC inhibited the hypoxia-induced increase in cytosolic Ca2+ concentration ([Ca2+]i). Co-immunoprecipitation and proximity ligation assays revealed increased interaction of PTEN with TRPC6 in human PASMC and murine lungs in response to hypoxia. In hypoxic PASMC, both PTEN and TRPC6 translocated to caveolae, and this response was blocked by pharmacological inhibition of Rho-associated protein kinase (ROCK) which in parallel prevented PTEN-TRPC6 interaction, hypoxia-induced [Ca2+]i increase, and HPV in PASMC and murine lungs, respectively.
Conclusion: Our data indicate a novel interplay between ROCK and [Ca2+]i signalling in HPV via PTEN, in that ROCK mediates interaction of PTEN and TRPC6 which then conjointly translocate to caveolae allowing for Ca2+ influx into and subsequent contraction of PASMC.
Keywords: Hypoxia; Phosphatase and tensin homolog (PTEN); Pulmonary artery smooth muscle cells (PASMC); Rho kinase (ROCK); Transient receptor potential canonical 6 (TRPC6).
Published on behalf of the European Society of Cardiology. All rights reserved. © The Author 2017. For permissions, please email: journals.permissions@oup.com.
Figures

Similar articles
-
Pannexin 1: a novel regulator of acute hypoxic pulmonary vasoconstriction.Cardiovasc Res. 2022 Aug 24;118(11):2535-2547. doi: 10.1093/cvr/cvab326. Cardiovasc Res. 2022. PMID: 34668529 Free PMC article.
-
Notch Activation of Ca(2+) Signaling in the Development of Hypoxic Pulmonary Vasoconstriction and Pulmonary Hypertension.Am J Respir Cell Mol Biol. 2015 Sep;53(3):355-67. doi: 10.1165/rcmb.2014-0235OC. Am J Respir Cell Mol Biol. 2015. PMID: 25569851 Free PMC article.
-
CFTR and sphingolipids mediate hypoxic pulmonary vasoconstriction.Proc Natl Acad Sci U S A. 2015 Mar 31;112(13):E1614-23. doi: 10.1073/pnas.1421190112. Epub 2015 Mar 17. Proc Natl Acad Sci U S A. 2015. PMID: 25829545 Free PMC article.
-
Hypoxic pulmonary vasoconstriction: redox regulation of O2-sensitive K+ channels by a mitochondrial O2-sensor in resistance artery smooth muscle cells.J Mol Cell Cardiol. 2004 Dec;37(6):1119-36. doi: 10.1016/j.yjmcc.2004.09.007. J Mol Cell Cardiol. 2004. PMID: 15572043 Review.
-
Molecular identification of O2 sensors and O2-sensitive potassium channels in the pulmonary circulation.Adv Exp Med Biol. 2000;475:219-40. doi: 10.1007/0-306-46825-5_21. Adv Exp Med Biol. 2000. PMID: 10849663 Review.
Cited by
-
Fasudil Dichloroacetate Alleviates SU5416/Hypoxia-Induced Pulmonary Arterial Hypertension by Ameliorating Dysfunction of Pulmonary Arterial Smooth Muscle Cells.Drug Des Devel Ther. 2021 Apr 22;15:1653-1666. doi: 10.2147/DDDT.S297500. eCollection 2021. Drug Des Devel Ther. 2021. PMID: 33935492 Free PMC article.
-
Pannexin 1: a novel regulator of acute hypoxic pulmonary vasoconstriction.Cardiovasc Res. 2022 Aug 24;118(11):2535-2547. doi: 10.1093/cvr/cvab326. Cardiovasc Res. 2022. PMID: 34668529 Free PMC article.
-
Hypoxia in Aging and Aging-Related Diseases: Mechanism and Therapeutic Strategies.Int J Mol Sci. 2022 Jul 25;23(15):8165. doi: 10.3390/ijms23158165. Int J Mol Sci. 2022. PMID: 35897741 Free PMC article. Review.
-
Rho kinase signaling and cardiac physiology.Curr Opin Physiol. 2018 Feb;1:14-20. doi: 10.1016/j.cophys.2017.07.005. Epub 2017 Dec 13. Curr Opin Physiol. 2018. PMID: 29527586 Free PMC article.
-
Effects of the Rho/Rho-Kinase Pathway on Perfusion Pressure in the Isolated-Perfused Rat Hind Limb Vascular Bed.Balkan Med J. 2021 Sep;38(5):304-309. doi: 10.5152/balkanmedj.2021.20014. Balkan Med J. 2021. PMID: 34558416 Free PMC article.
References
-
- Marshall BE, Hanson CW, Frasch F, Marshall C.. Role of hypoxic pulmonary vasoconstriction in pulmonary gas exchange and blood flow distribution. 2. Pathophysiol Intens Care Med 1994;20:379–389. - PubMed
-
- McCormack DG, Paterson NAM.. Loss of hypoxic pulmonary vasoconstriction in chronic pneumonia is not mediated by nitric oxide. Am J Physiol 1993; 265:H1523–H1528. - PubMed
-
- Sommer N, Dietrich A, Schermuly RT, Ghofrani HA, Gudermann T, Schulz R, Seeger W, Grimminger F, Weissmann N.. Regulation of hypoxic pulmonary vasoconstriction: basic mechanisms. Eur Respir J 2008;32:1639–1651. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
