

Phagocytosis imprints heterogeneity in tissue-resident macrophages
- PMID: 28432199
- PMCID: PMC5413334
- DOI: 10.1084/jem.20161375
Phagocytosis imprints heterogeneity in tissue-resident macrophages
Abstract
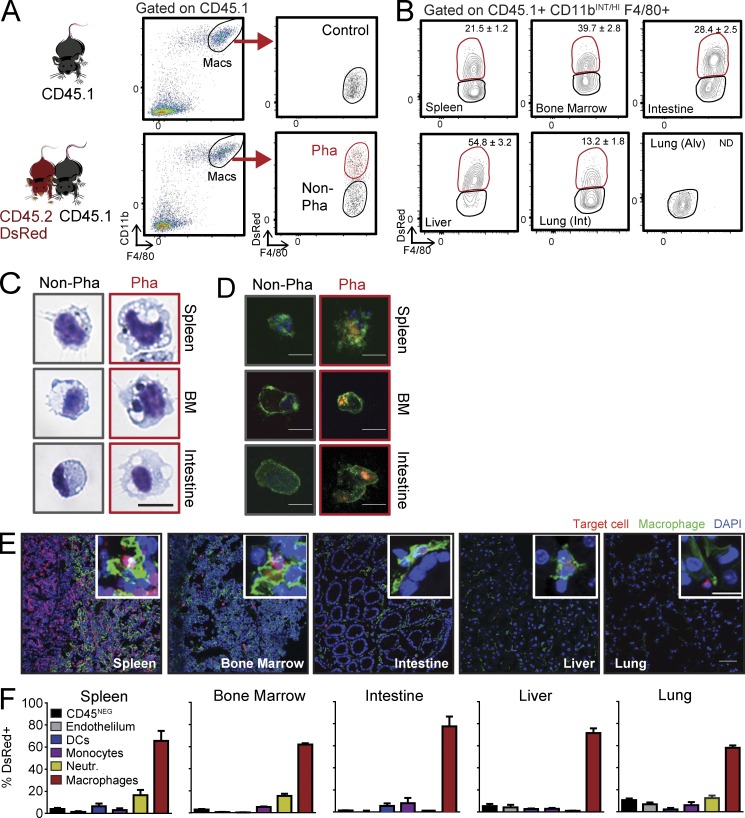
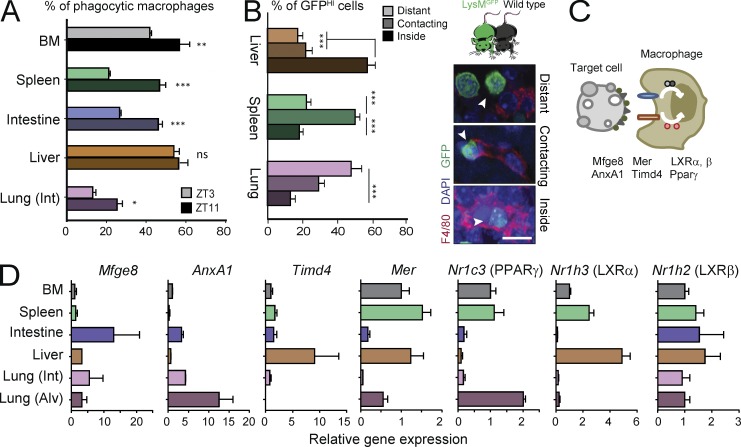
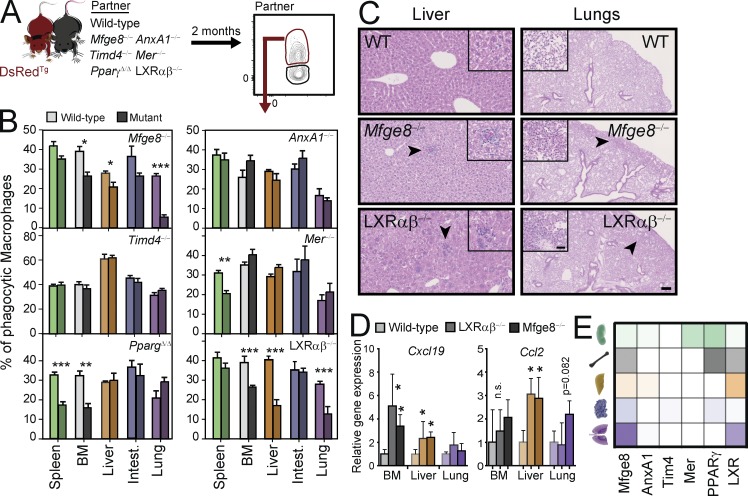
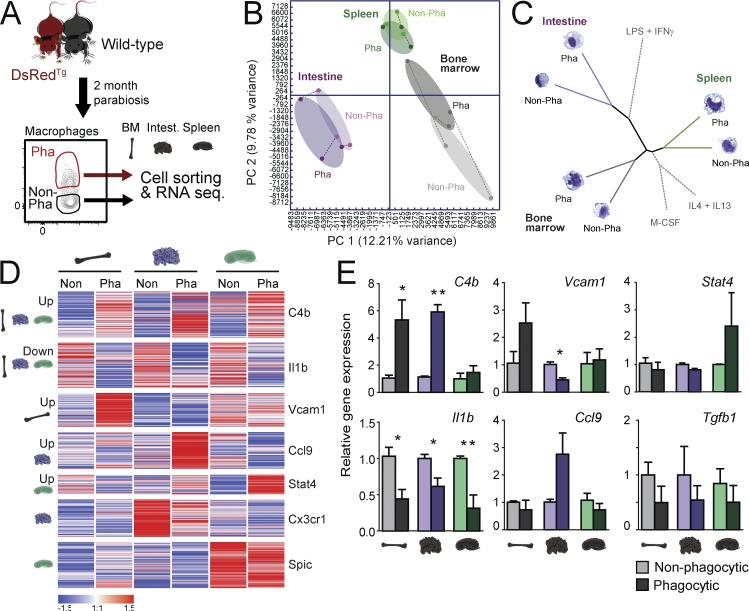
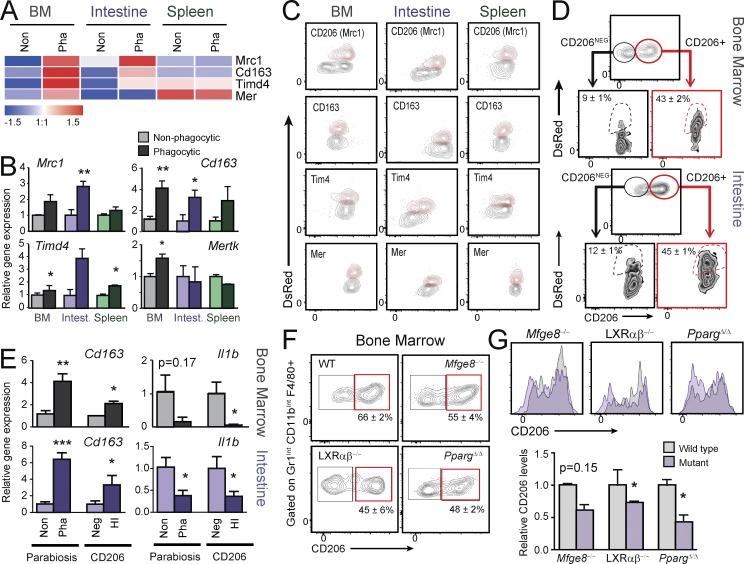
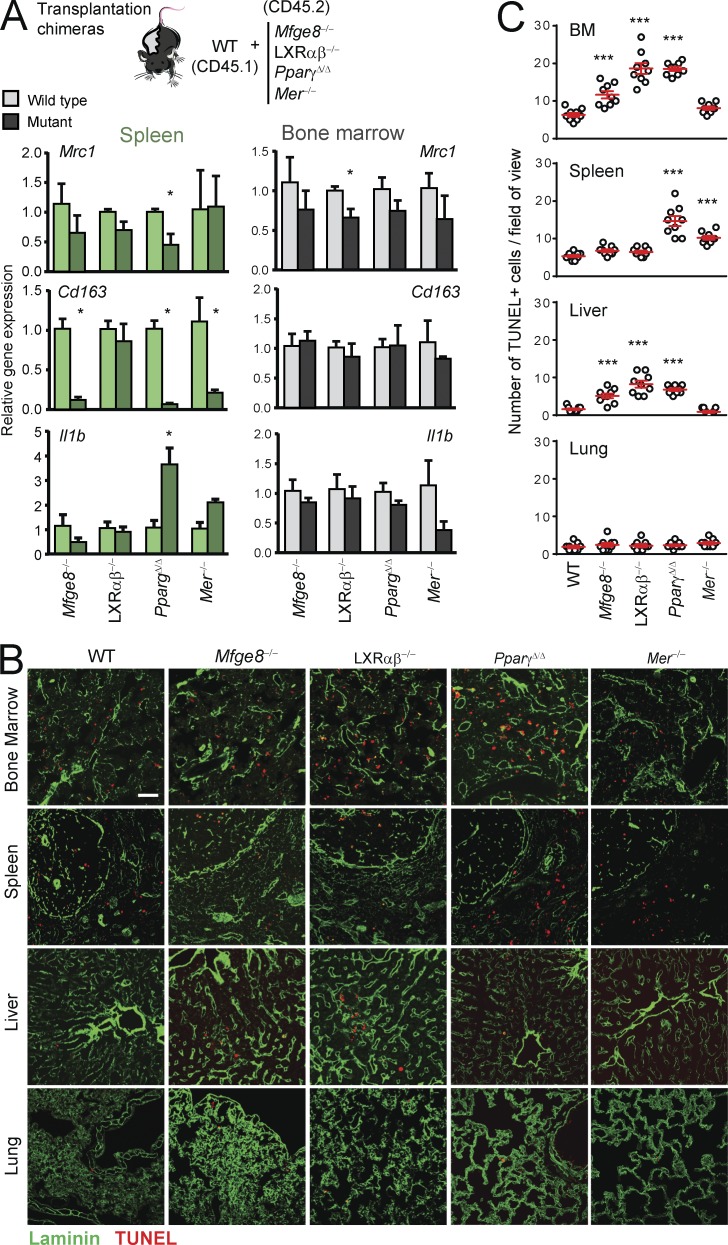
Tissue-resident macrophages display varying phenotypic and functional properties that are largely specified by their local environment. One of these functions, phagocytosis, mediates the natural disposal of billions of cells, but its mechanisms and consequences within living tissues are poorly defined. Using a parabiosis-based strategy, we identified and isolated macrophages from multiple tissues as they phagocytosed blood-borne cellular material. Phagocytosis was circadianally regulated and mediated by distinct repertoires of receptors, opsonins, and transcription factors in macrophages from each tissue. Although the tissue of residence defined the core signature of macrophages, phagocytosis imprinted a distinct antiinflammatory profile. Phagocytic macrophages expressed CD206, displayed blunted expression of Il1b, and supported tissue homeostasis. Thus, phagocytosis is a source of macrophage heterogeneity that acts together with tissue-derived factors to preserve homeostasis.
© 2017 A-Gonzalez et al.
Figures
References
-
- A-Gonzalez N., Bensinger S.J., Hong C., Beceiro S., Bradley M.N., Zelcer N., Deniz J., Ramirez C., Díaz M., Gallardo G., et al. 2009. Apoptotic cells promote their own clearance and immune tolerance through activation of the nuclear receptor LXR. Immunity. 31:245–258. 10.1016/j.immuni.2009.06.018 - DOI - PMC - PubMed
-
- A-Gonzalez N., Guillen J.A., Gallardo G., Diaz M., de la Rosa J.V., Hernandez I.H., Casanova-Acebes M., Lopez F., Tabraue C., Beceiro S., et al. 2013. The nuclear receptor LXRα controls the functional specialization of splenic macrophages. Nat. Immunol. 14:831–839. 10.1038/ni.2622 - DOI - PMC - PubMed
-
- Bosurgi L., Bernink J.H., Delgado Cuevas V., Gagliani N., Joannas L., Schmid E.T., Booth C.J., Ghosh S., and Rothlin C.V.. 2013. Paradoxical role of the proto-oncogene Axl and Mer receptor tyrosine kinases in colon cancer. Proc. Natl. Acad. Sci. USA. 110:13091–13096. 10.1073/pnas.1302507110 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
