

The U2AF1S34F mutation induces lineage-specific splicing alterations in myelodysplastic syndromes
- PMID: 28436936
- PMCID: PMC5451246
- DOI: 10.1172/JCI91363
The U2AF1S34F mutation induces lineage-specific splicing alterations in myelodysplastic syndromes
Erratum in
-
The U2AF1S34F mutation induces lineage-specific splicing alterations in myelodysplastic syndromes.J Clin Invest. 2017 Sep 1;127(9):3557. doi: 10.1172/JCI96202. Epub 2017 Sep 1. J Clin Invest. 2017. PMID: 28862641 Free PMC article. No abstract available.
Abstract
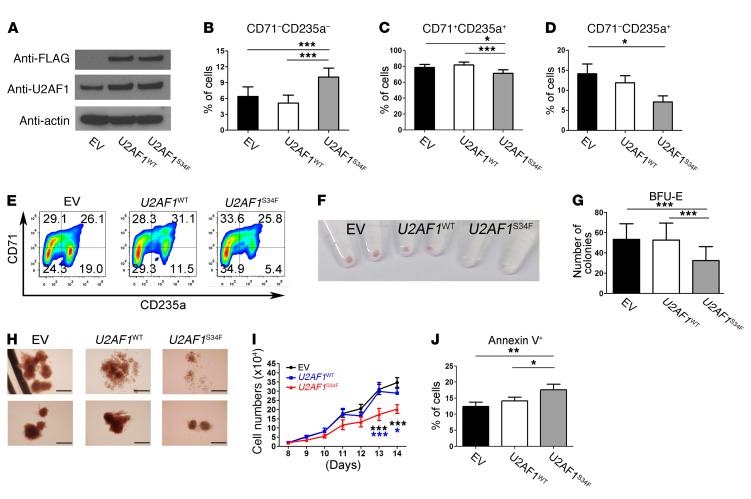
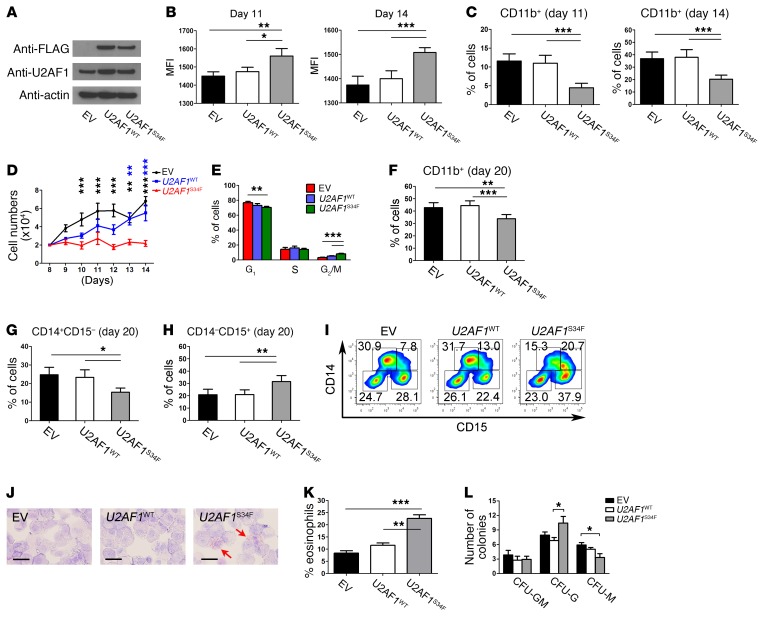
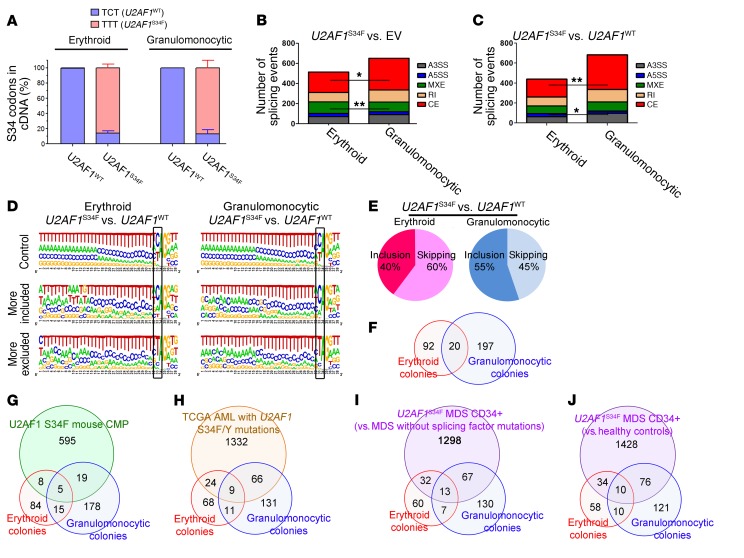
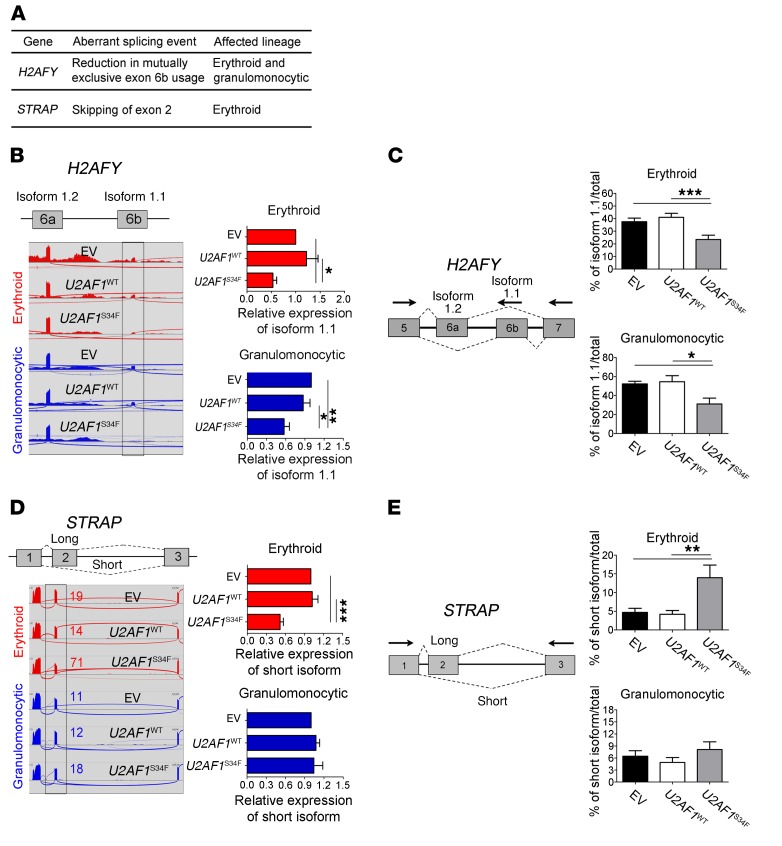
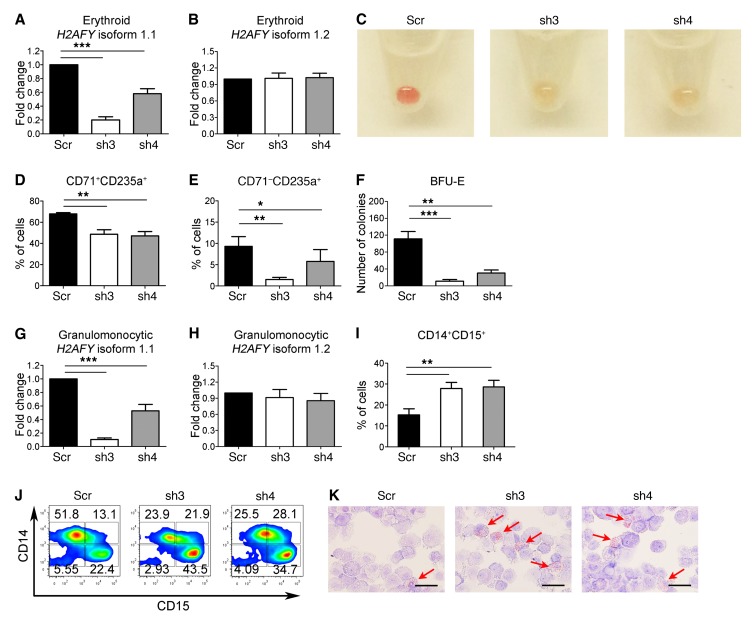
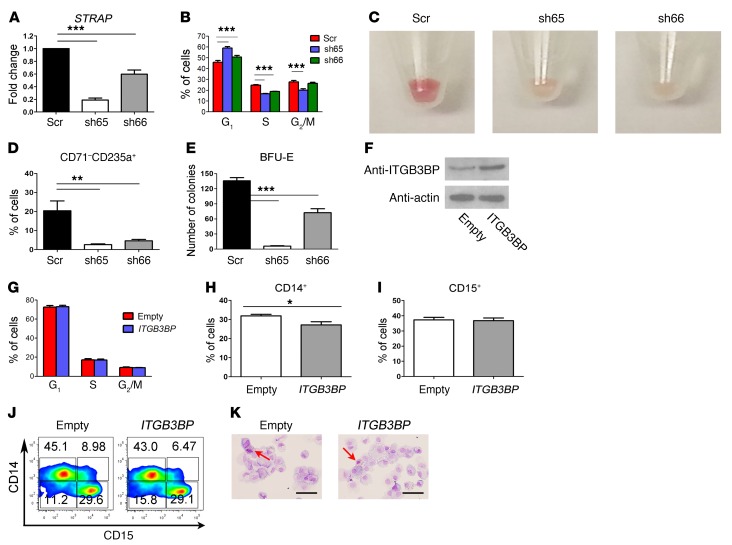
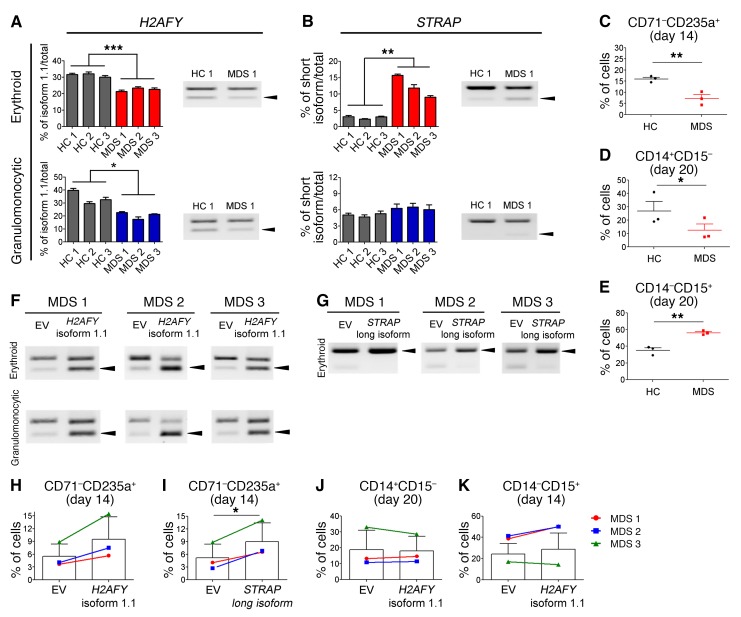
Mutations of the splicing factor-encoding gene U2AF1 are frequent in the myelodysplastic syndromes (MDS), a myeloid malignancy, and other cancers. Patients with MDS suffer from peripheral blood cytopenias, including anemia, and an increasing percentage of bone marrow myeloblasts. We studied the impact of the common U2AF1S34F mutation on cellular function and mRNA splicing in the main cell lineages affected in MDS. We demonstrated that U2AF1S34F expression in human hematopoietic progenitors impairs erythroid differentiation and skews granulomonocytic differentiation toward granulocytes. RNA sequencing of erythroid and granulomonocytic colonies revealed that U2AF1S34F induced a higher number of cassette exon splicing events in granulomonocytic cells than in erythroid cells. U2AF1S34F altered mRNA splicing of many transcripts that were expressed in both cell types in a lineage-specific manner. In hematopoietic progenitors, the introduction of isoform changes identified in the U2AF1S34F target genes H2AFY, encoding an H2A histone variant, and STRAP, encoding serine/threonine kinase receptor-associated protein, recapitulated phenotypes associated with U2AF1S34F expression in erythroid and granulomonocytic cells, suggesting a causal link. Furthermore, we showed that isoform modulation of H2AFY and STRAP rescues the erythroid differentiation defect in U2AF1S34F MDS cells, suggesting that splicing modulators could be used therapeutically. These data have critical implications for understanding MDS phenotypic heterogeneity and support the development of therapies targeting splicing abnormalities.
Conflict of interest statement
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
