

Desmoglein 2 modulates extracellular vesicle release from squamous cell carcinoma keratinocytes
- PMID: 28438789
- PMCID: PMC5503718
- DOI: 10.1096/fj.201601138RR
Desmoglein 2 modulates extracellular vesicle release from squamous cell carcinoma keratinocytes
Abstract
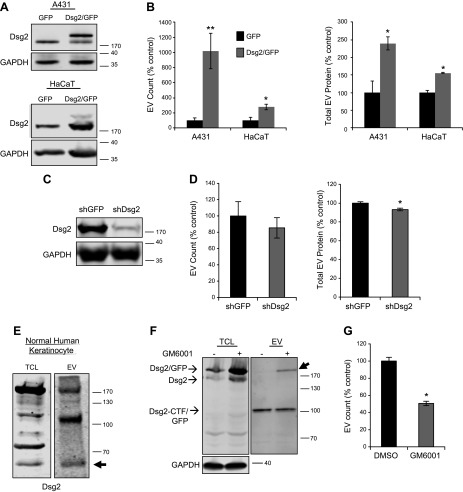
Extracellular vesicles (EVs) are nanoscale membrane-derived vesicles that serve as intercellular messengers carrying lipids, proteins, and genetic material. Substantial evidence has shown that cancer-derived EVs, secreted by tumor cells into the blood and other bodily fluids, play a critical role in modulating the tumor microenvironment and affecting the pathogenesis of cancer. Here we demonstrate for the first time that squamous cell carcinoma (SCC) EVs were enriched with the C-terminal fragment of desmoglein 2 (Dsg2), a desmosomal cadherin often overexpressed in malignancies. Overexpression of Dsg2 increased EV release and mitogenic content including epidermal growth factor receptor and c-Src. Inhibiting ectodomain shedding of Dsg2 with the matrix metalloproteinase inhibitor GM6001 resulted in accumulation of full-length Dsg2 in EVs and reduced EV release. When cocultured with Dsg2/green fluorescence protein-expressing SCC cells, green fluorescence protein signal was detected by fluorescence-activated cell sorting analysis in the CD90+ fibroblasts. Furthermore, SCC EVs activated Erk1/2 and Akt signaling and enhanced fibroblast cell proliferation. In vivo, Dsg2 was highly up-regulated in the head and neck SCCs, and EVs isolated from sera of patients with SCC were enriched in Dsg2 C-terminal fragment and epidermal growth factor receptor. This study defines a mechanism by which Dsg2 expression in cancer cells can modulate the tumor microenvironment, a step critical for tumor progression.-Overmiller, A. M., Pierluissi, J. A., Wermuth, P. J., Sauma, S., Martinez-Outschoorn, U., Tuluc, M., Luginbuhl, A., Curry, J., Harshyne, L. A., Wahl, J. K. III, South, A. P., Mahoney, M. G. Desmoglein 2 modulates extracellular vesicle release from squamous cell carcinoma keratinocytes.
Keywords: EGFR; HNSCC; exosome; fibroblast.
© FASEB.
Figures






Similar articles
-
miRNA- and cytokine-associated extracellular vesicles mediate squamous cell carcinomas.J Extracell Vesicles. 2020 Jul 13;9(1):1790159. doi: 10.1080/20013078.2020.1790159. J Extracell Vesicles. 2020. PMID: 32944178 Free PMC article.
-
c-Src/Cav1-dependent activation of the EGFR by Dsg2.Oncotarget. 2016 Jun 21;7(25):37536-37555. doi: 10.18632/oncotarget.7675. Oncotarget. 2016. PMID: 26918609 Free PMC article.
-
A molecular study of desmosomes identifies a desmoglein isoform switch in head and neck squamous cell carcinoma.J Oral Pathol Med. 2011 Jan;40(1):67-76. doi: 10.1111/j.1600-0714.2010.00951.x. Epub 2010 Oct 4. J Oral Pathol Med. 2011. PMID: 20923451
-
Characteristics and roles of extracellular vesicles released by epidermal keratinocytes.J Eur Acad Dermatol Venereol. 2019 Dec;33(12):2264-2272. doi: 10.1111/jdv.15859. Epub 2019 Sep 9. J Eur Acad Dermatol Venereol. 2019. PMID: 31403744 Review.
-
Extracellular vesicle communication pathways as regulatory targets of oncogenic transformation.Semin Cell Dev Biol. 2017 Jul;67:11-22. doi: 10.1016/j.semcdb.2017.01.003. Epub 2017 Jan 8. Semin Cell Dev Biol. 2017. PMID: 28077296 Review.
Cited by
-
Enhancement of Cutaneous Wound Healing by Dsg2 Augmentation of uPAR Secretion.J Invest Dermatol. 2018 Nov;138(11):2470-2479. doi: 10.1016/j.jid.2018.04.024. Epub 2018 May 9. J Invest Dermatol. 2018. PMID: 29753032 Free PMC article.
-
The bronchoalveolar lavage fluid CD44 as a marker for pulmonary fibrosis in diffuse parenchymal lung diseases.Front Immunol. 2025 Jan 13;15:1479458. doi: 10.3389/fimmu.2024.1479458. eCollection 2024. Front Immunol. 2025. PMID: 39872532 Free PMC article.
-
Exosomes and cancer - Diagnostic and prognostic biomarkers and therapeutic vehicle.Oncogenesis. 2022 Sep 15;11(1):54. doi: 10.1038/s41389-022-00431-5. Oncogenesis. 2022. PMID: 36109501 Free PMC article. Review.
-
Desmoglein-2 is important for islet function and β-cell survival.Cell Death Dis. 2022 Oct 29;13(10):911. doi: 10.1038/s41419-022-05326-2. Cell Death Dis. 2022. PMID: 36309486 Free PMC article.
-
Reproduction of the Antitumor Effect of Cisplatin and Cetuximab Using a Three-dimensional Spheroid Model in Oral Cancer.Int J Med Sci. 2022 Jul 18;19(8):1320-1333. doi: 10.7150/ijms.74109. eCollection 2022. Int J Med Sci. 2022. PMID: 35928727 Free PMC article.
References
-
- Colombo M., Raposo G., Théry C. (2014) Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu. Rev. Cell Dev. Biol. 30, 255–289 - PubMed
-
- Hood J. L., San R. S., Wickline S. A. (2011) Exosomes released by melanoma cells prepare sentinel lymph nodes for tumor metastasis. Cancer Res. 71, 3792–3801 - PubMed
-
- Peinado H., Alečković M., Lavotshkin S., Matei I., Costa-Silva B., Moreno-Bueno G., Hergueta-Redondo M., Williams C., García-Santos G., Ghajar C., Nitadori-Hoshino A., Hoffman C., Badal K., Garcia B. A., Callahan M. K., Yuan J., Martins V. R., Skog J., Kaplan R. N., Brady M. S., Wolchok J. D., Chapman P. B., Kang Y., Bromberg J., Lyden D. (2012) Melanoma exosomes educate bone marrow progenitor cells toward a pro-metastatic phenotype through MET. Nat. Med. 18, 883–891 - PMC - PubMed
-
- Jemal A., Bray F., Center M. M., Ferlay J., Ward E., Forman D. (2011) Global cancer statistics. CA Cancer J. Clin. 61, 69–90 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
