

Allosteric mechanism of action of the therapeutic anti-IgE antibody omalizumab
- PMID: 28438838
- PMCID: PMC5473249
- DOI: 10.1074/jbc.M117.776476
Allosteric mechanism of action of the therapeutic anti-IgE antibody omalizumab
Abstract
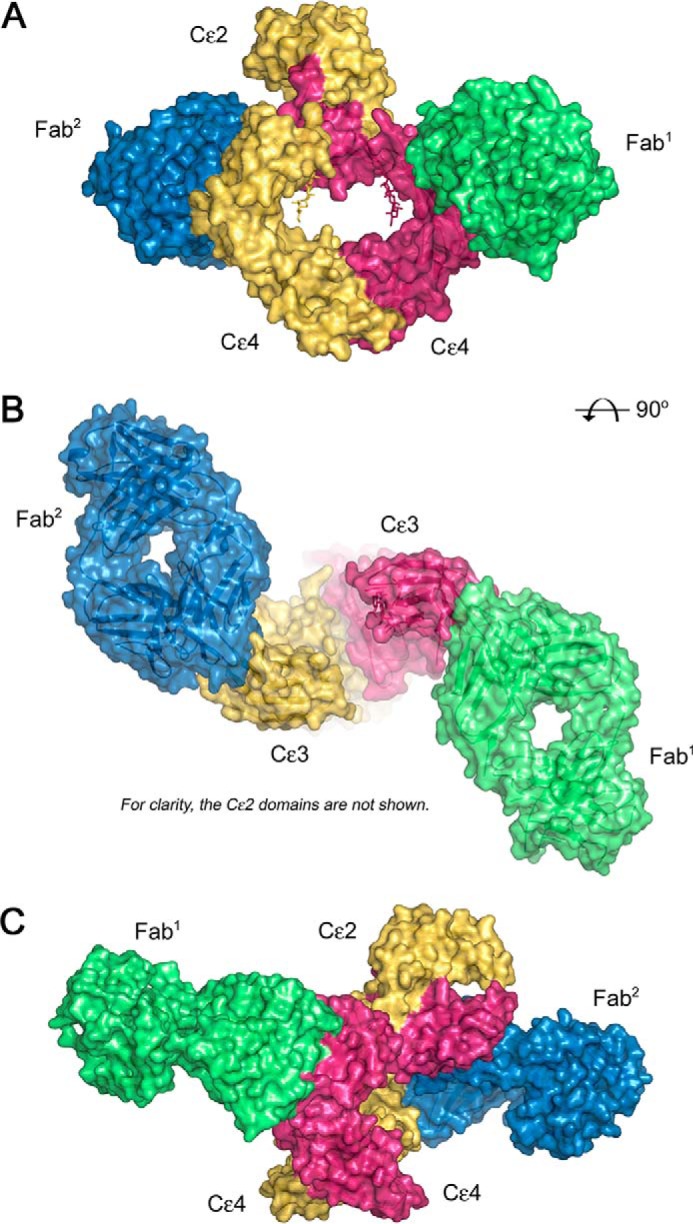
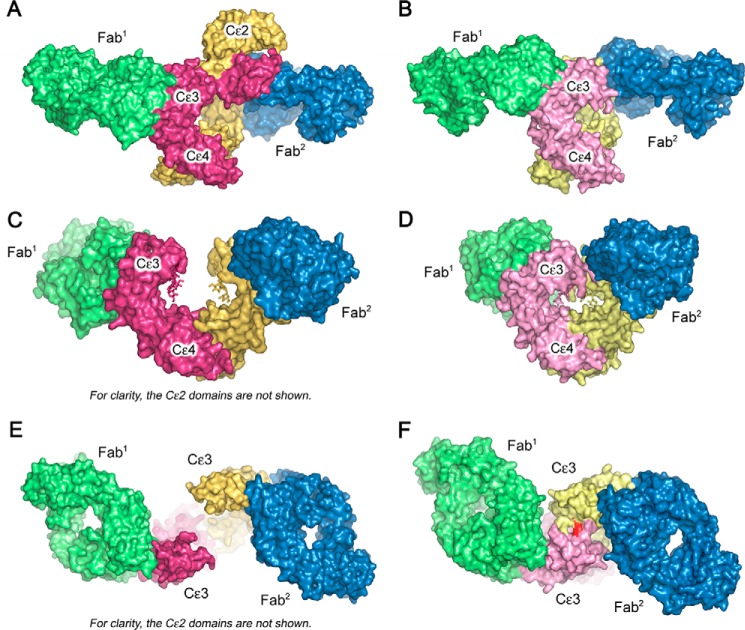
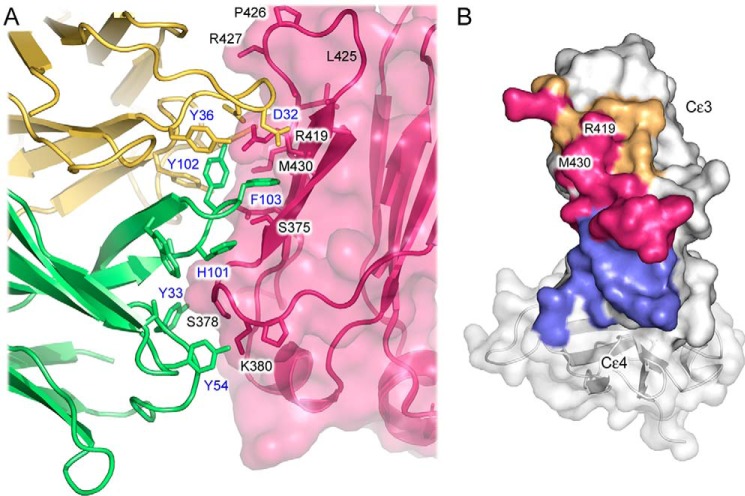
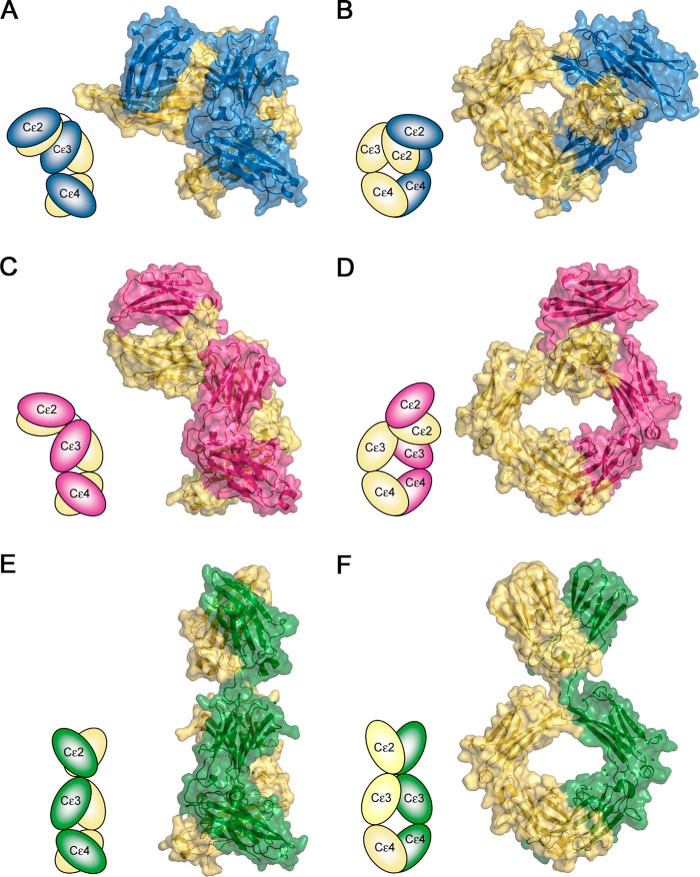
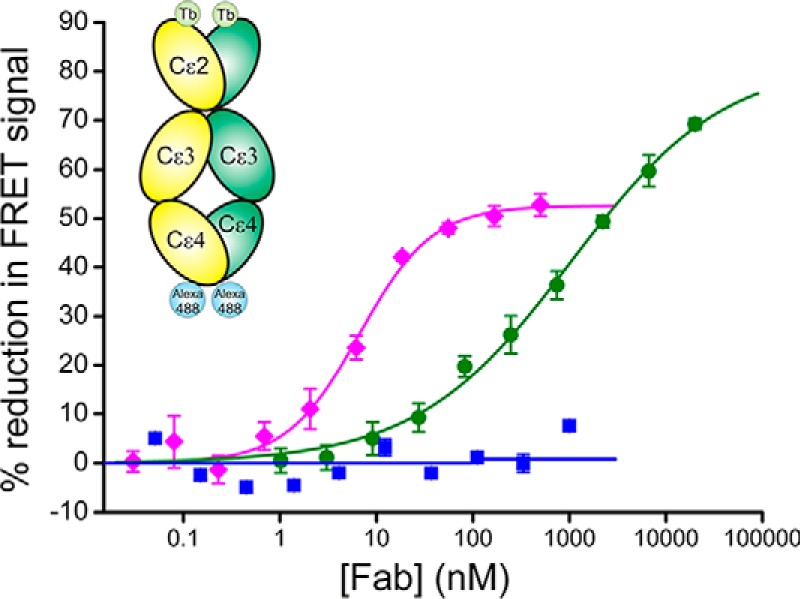
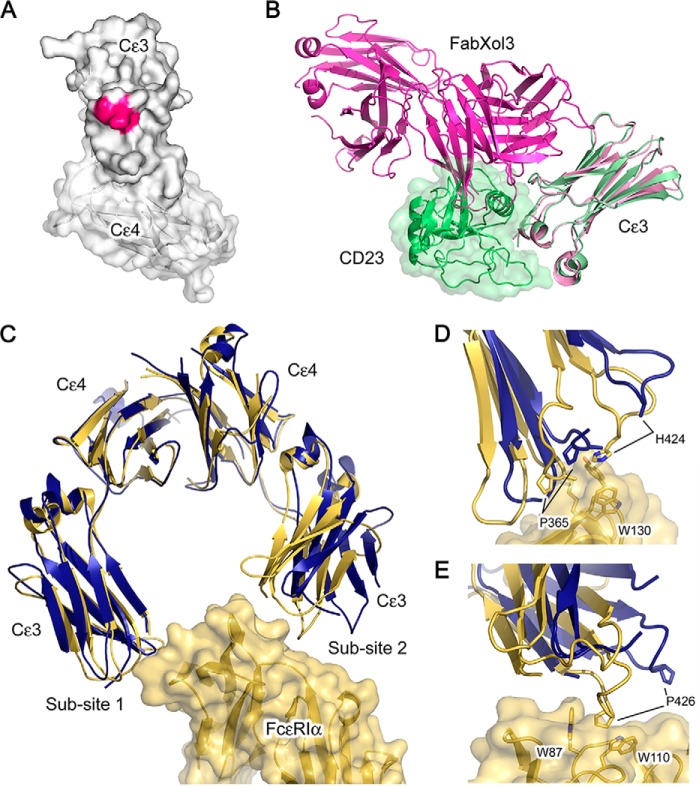
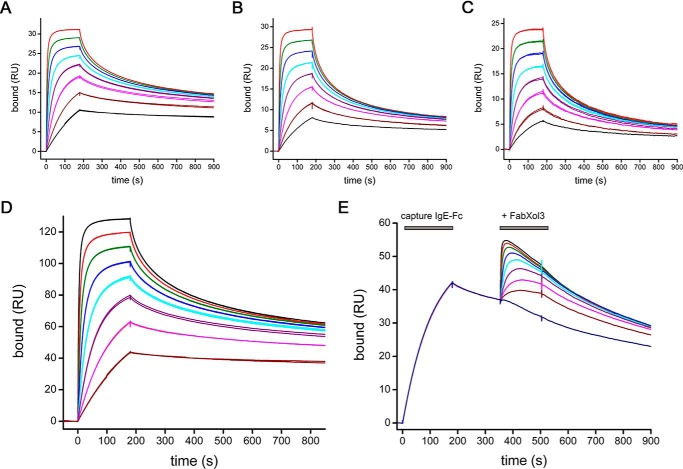
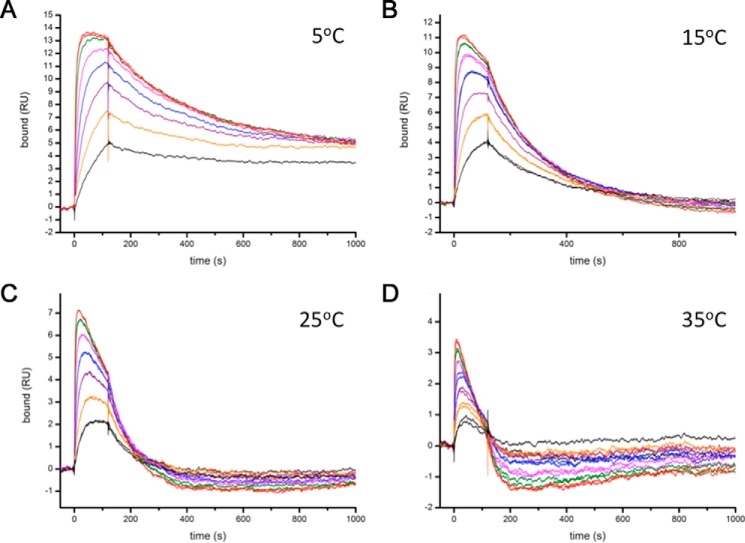
Immunoglobulin E and its interactions with receptors FcϵRI and CD23 play a central role in allergic disease. Omalizumab, a clinically approved therapeutic antibody, inhibits the interaction between IgE and FcϵRI, preventing mast cell and basophil activation, and blocks IgE binding to CD23 on B cells and antigen-presenting cells. We solved the crystal structure of the complex between an omalizumab-derived Fab and IgE-Fc, with one Fab bound to each Cϵ3 domain. Free IgE-Fc adopts an acutely bent structure, but in the complex it is only partially bent, with large-scale conformational changes in the Cϵ3 domains that inhibit the interaction with FcϵRI. CD23 binding is inhibited sterically due to overlapping binding sites on each Cϵ3 domain. Studies of omalizumab Fab binding in solution demonstrate the allosteric basis for FcϵRI inhibition and, together with the structure, reveal how omalizumab may accelerate dissociation of receptor-bound IgE from FcϵRI, exploiting the intrinsic flexibility and allosteric potential of IgE.
Keywords: X-ray crystallography; allergy; allosteric regulation; antibody; immunoglobulin E (IgE); omalizumab.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
J. D., B. P. C., T. C., G. C., M. W., and A. J. H. are employees of UCB-Celltech. T. C., G. C., and A. J. H. are in receipt of stock options. UCB-Celltech has filed a patent application based on elements of this work with the UK Intellectual Property Office
Figures
References
-
- Gould H. J., and Sutton B. J. (2008) IgE in allergy and asthma today. Nat. Rev. Immunol. 8, 205–217 - PubMed
-
- Sutton B. J., and Davies A. M. (2015) Structure and dynamics of IgE–receptor interactions: FcϵRI and CD23/FcϵRII. Immunol. Rev. 268, 222–235 - PubMed
-
- Holowka D., and Baird B. (1983) Structural studies on the membrane-bound immunoglobulin E (IgE)-receptor complex. 2. Mapping of distances between sites on IgE and the membrane surface. Biochemistry 22, 3475–3484 - PubMed
-
- Zheng Y., Shopes B., Holowka D., and Baird B. (1991) Conformations of IgE bound to its receptor FcϵRI and in solution. Biochemistry 30, 9125–9132 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
