

Novel clades of the HU/IHF superfamily point to unexpected roles in the eukaryotic centrosome, chromosome partitioning, and biologic conflicts
- PMID: 28441108
- PMCID: PMC5499826
- DOI: 10.1080/15384101.2017.1315494
Novel clades of the HU/IHF superfamily point to unexpected roles in the eukaryotic centrosome, chromosome partitioning, and biologic conflicts
Abstract
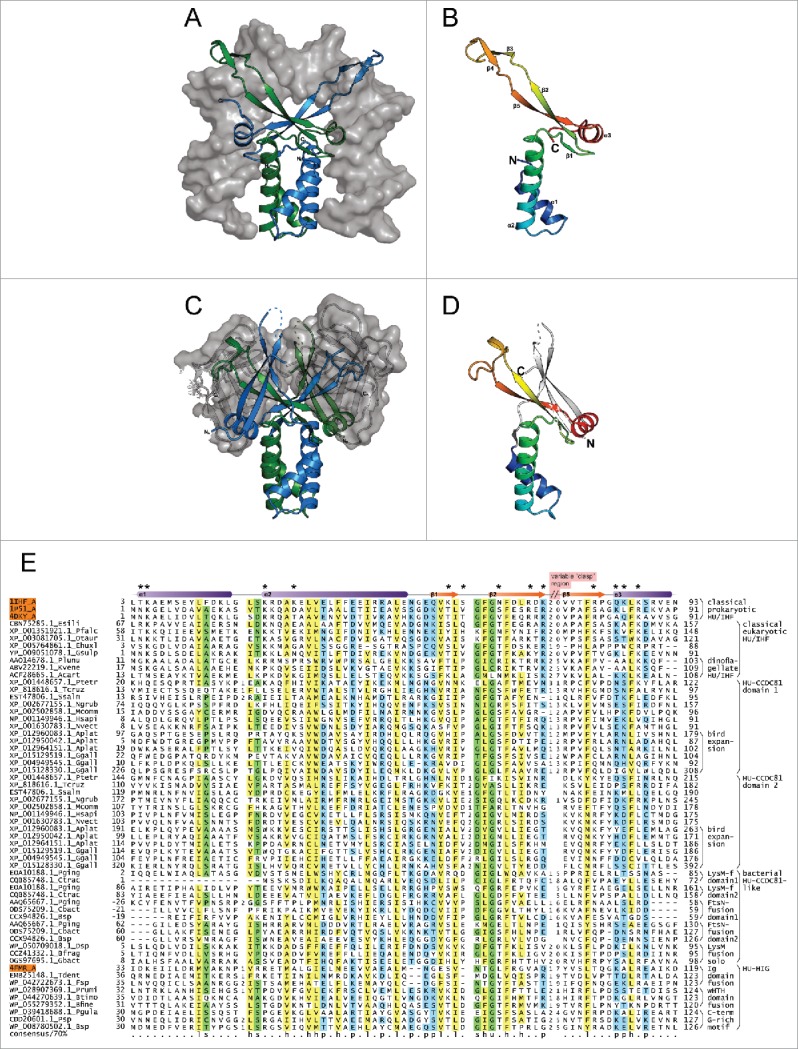
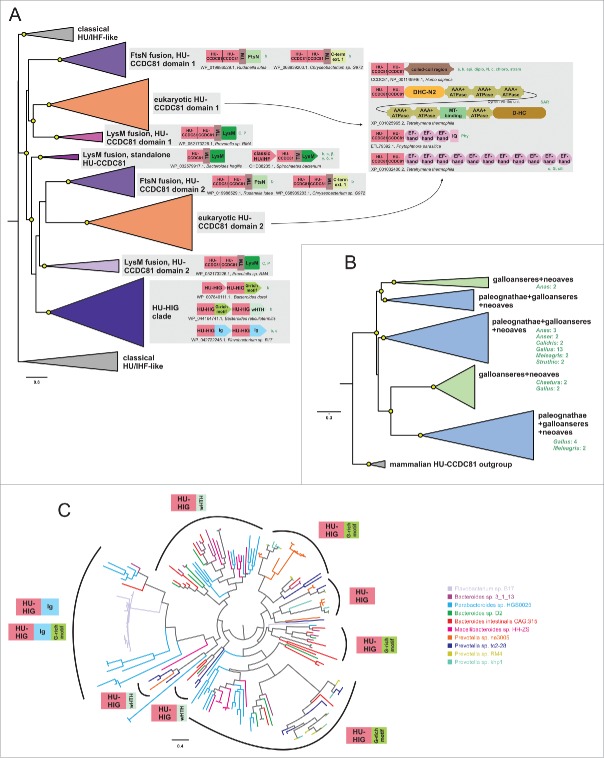
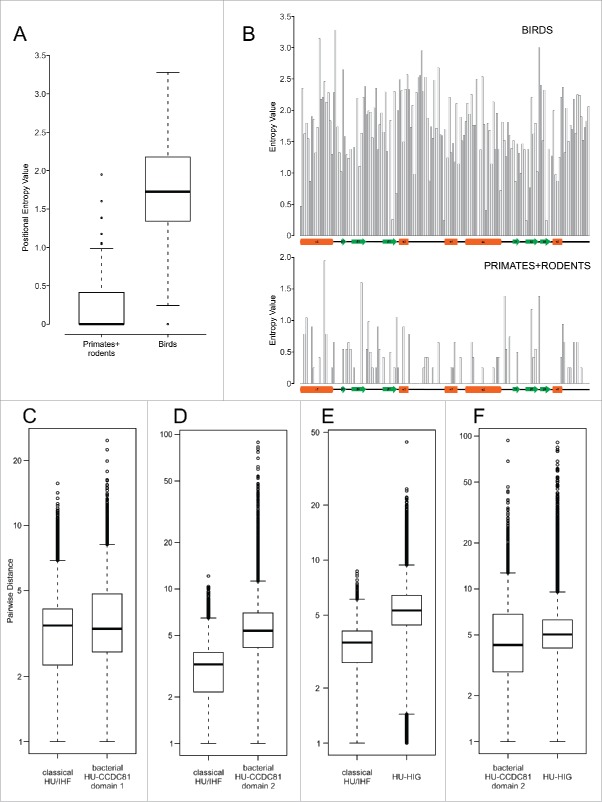
The HU superfamily of proteins, with a unique DNA-binding mode, has been extensively studied as the primary chromosome-packaging protein of the bacterial superkingdom. Representatives also play a role in DNA-structuring during recombination events and in eukaryotic organellar genome maintenance. However, beyond these well-studied roles, little is understood of the functional diversification of this large superfamily. Using sensitive sequence and structure analysis methods we identify multiple novel clades of the HU superfamily. We present evidence that a novel eukaryotic clade prototyped by the human CCDC81 protein acquired roles beyond DNA-binding, likely in protein-protein interaction in centrosome organization and as a potential cargo-binding protein in conjunction with Dynein-VII. We also show that these eukaryotic versions were acquired via an early lateral transfer from bacteroidetes, where we predict a role in chromosome partition. This likely happened before the last eukaryotic common ancestor, pointing to potential endosymbiont contributions beyond that of the mitochondrial progenitor. Further, we show that the dramatic lineage-specific expansion of this domain in the bacteroidetes lineage primarily is linked to a functional shift related to potential recognition and preemption of genome invasive entities such as mobile elements. Remarkably, the CCDC81 clade has undergone a similar massive lineage-specific expansion within the archosaurian lineage in birds, suggesting a possible use of the HU superfamily in a similar capacity in recognition of non-self molecules even in this case.
Keywords: CCDC81; HU; IHF; bacteroidetes; biological conflict; birds; centrosome.
Figures
Similar articles
-
Identification of novel families and classification of the C2 domain superfamily elucidate the origin and evolution of membrane targeting activities in eukaryotes.Gene. 2010 Dec 1;469(1-2):18-30. doi: 10.1016/j.gene.2010.08.006. Epub 2010 Aug 14. Gene. 2010. PMID: 20713135 Free PMC article.
-
Structural and evolutionary analyses reveal determinants of DNA binding specificities of nucleoid-associated proteins HU and IHF.Mol Phylogenet Evol. 2017 Feb;107:356-366. doi: 10.1016/j.ympev.2016.11.014. Epub 2016 Nov 25. Mol Phylogenet Evol. 2017. PMID: 27894997
-
The Evolution of Sequence Specificity in a DNA Binding Protein Family.J Mol Biol. 2025 Jul 15;437(14):169177. doi: 10.1016/j.jmb.2025.169177. Epub 2025 Apr 29. J Mol Biol. 2025. PMID: 40311744
-
Histones, HMG, HU, IHF: Même combat.Biochimie. 1994;76(10-11):901-8. doi: 10.1016/0300-9084(94)90014-0. Biochimie. 1994. PMID: 7748933 Review.
-
RND transporters in the living world.Res Microbiol. 2018 Sep-Oct;169(7-8):363-371. doi: 10.1016/j.resmic.2018.03.001. Epub 2018 Mar 22. Res Microbiol. 2018. PMID: 29577985 Free PMC article. Review.
Cited by
-
Bacterial death and TRADD-N domains help define novel apoptosis and immunity mechanisms shared by prokaryotes and metazoans.Elife. 2021 Jun 1;10:e70394. doi: 10.7554/eLife.70394. Elife. 2021. PMID: 34061031 Free PMC article.
-
Shh agonist enhances maturation in homotypic Lgr5-positive inner ear organoids.Theranostics. 2025 Apr 13;15(12):5543-5565. doi: 10.7150/thno.107345. eCollection 2025. Theranostics. 2025. PMID: 40365278 Free PMC article.
-
Spatially distinct physiology of Bacteroides fragilis within the proximal colon of gnotobiotic mice.Nat Microbiol. 2020 May;5(5):746-756. doi: 10.1038/s41564-020-0683-3. Epub 2020 Mar 9. Nat Microbiol. 2020. PMID: 32152589 Free PMC article.
-
A novel mechanism of sperm midpiece epididymal maturation and the role of CCDC112 in sperm midpiece formation and establishing an optimal flagella waveform.Cell Commun Signal. 2025 Jul 1;23(1):319. doi: 10.1186/s12964-025-02320-x. Cell Commun Signal. 2025. PMID: 40598224 Free PMC article.
-
Functional Innovation in the Evolution of the Calcium-Dependent System of the Eukaryotic Endoplasmic Reticulum.Front Genet. 2020 Feb 6;11:34. doi: 10.3389/fgene.2020.00034. eCollection 2020. Front Genet. 2020. PMID: 32117448 Free PMC article.
References
-
- Dillon SC, Dorman CJ. Bacterial nucleoid-associated proteins, nucleoid structure and gene expression. Nat Rev Microbiol 2010; 8:185-95; PMID:20140026; https://doi.org/10.1038/nrmicro2261 - DOI - PubMed
-
- Reeve JN, Bailey KA, Li WT, Marc F, Sandman K, Soares DJ. Archaeal histones: structures, stability and DNA binding. Biochem Soc Trans 2004; 32:227-30; PMID:15046577; https://doi.org/10.1042/bst0320227 - DOI - PubMed
-
- Sandman K, Reeve JN. Archaeal chromatin proteins: different structures but common function? Curr Opin Microbiol 2005; 8:656-61; PMID:16256418; https://doi.org/10.1016/j.mib.2005.10.007 - DOI - PubMed
-
- Jones DO, Cowell IG, Singh PB. Mammalian chromodomain proteins: their role in genome organisation and expression. Bioessays 2000; 22:124-37; PMID:10655032; https://doi.org/10.1002/(SICI)1521-1878(200002)22:2%3c124::AID-BIES4%3e3... - DOI - PubMed
-
- Luijsterburg MS, White MF, van Driel R, Dame RT. The major architects of chromatin: architectural proteins in bacteria, archaea and eukaryotes. Crit Rev Biochem Mol Biol 2008; 43:393-418; PMID:19037758; https://doi.org/10.1080/10409230802528488 - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources