

Pax7 remodels the chromatin landscape in skeletal muscle stem cells
- PMID: 28441415
- PMCID: PMC5404880
- DOI: 10.1371/journal.pone.0176190
Pax7 remodels the chromatin landscape in skeletal muscle stem cells
Abstract
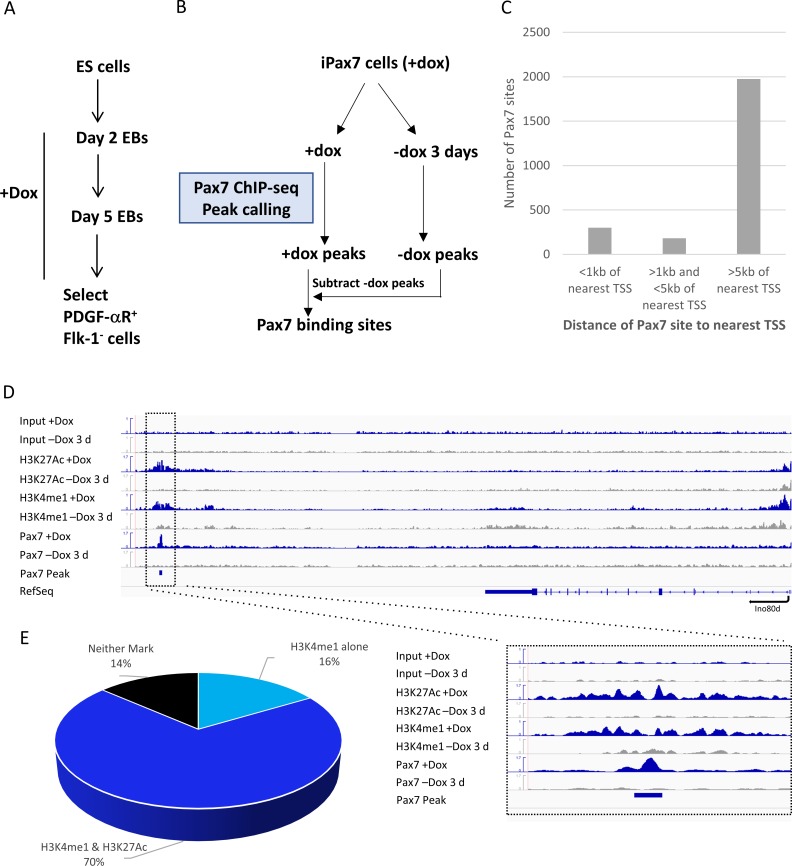
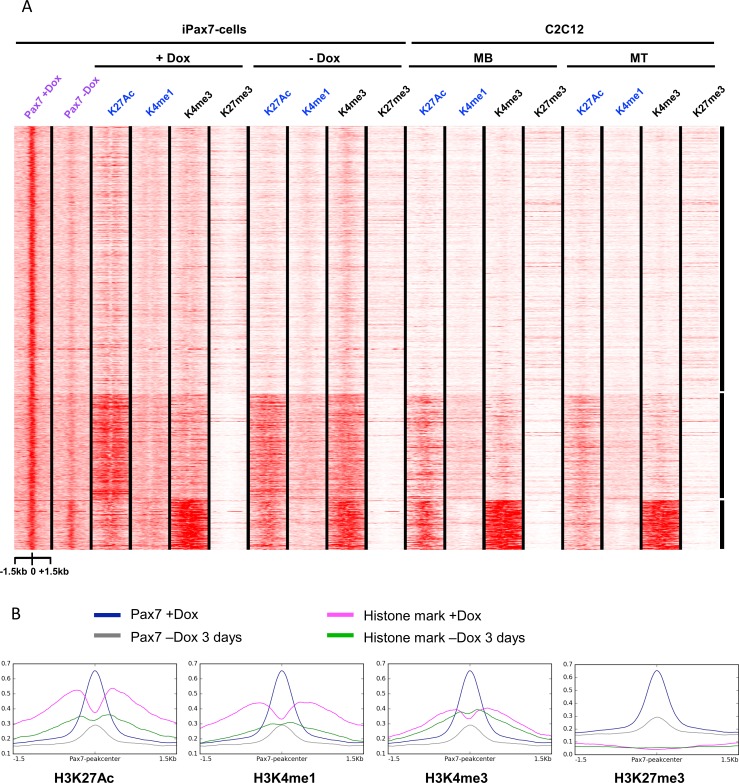
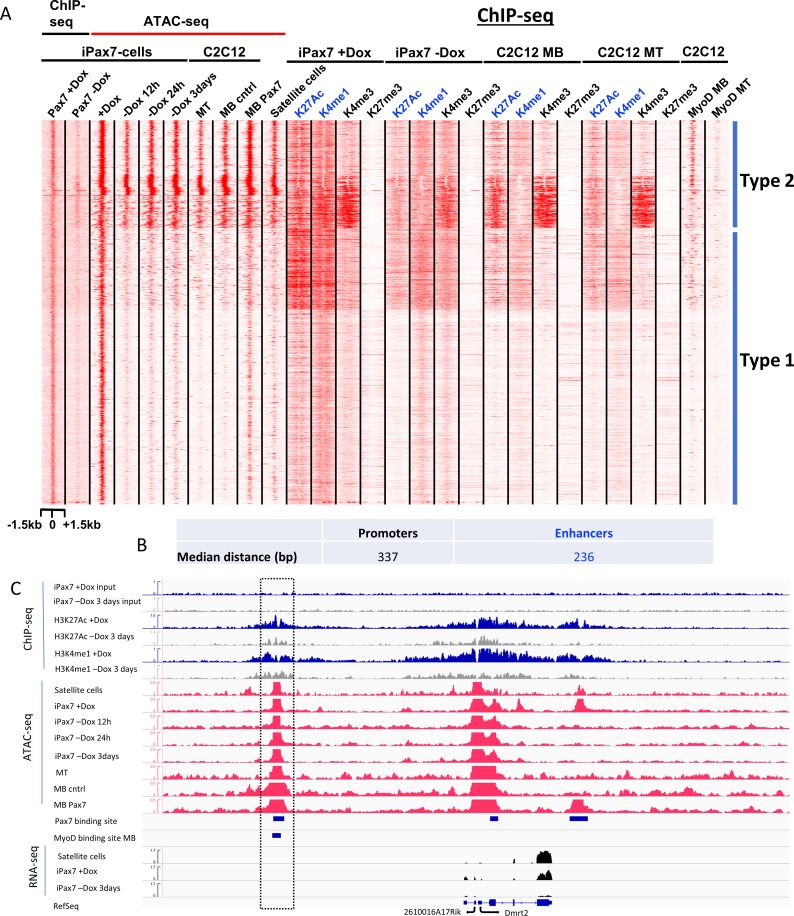
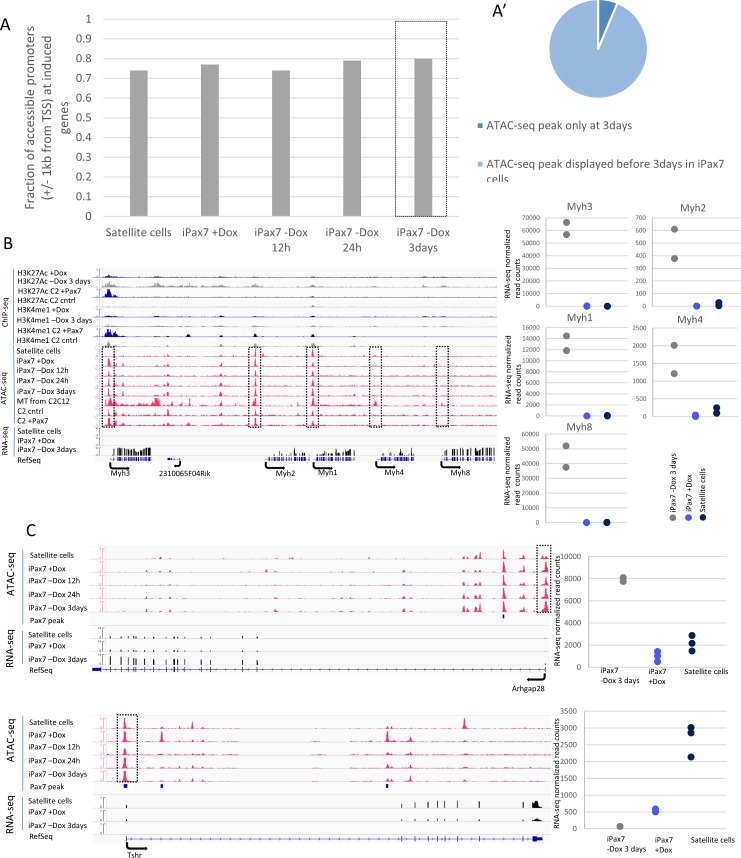
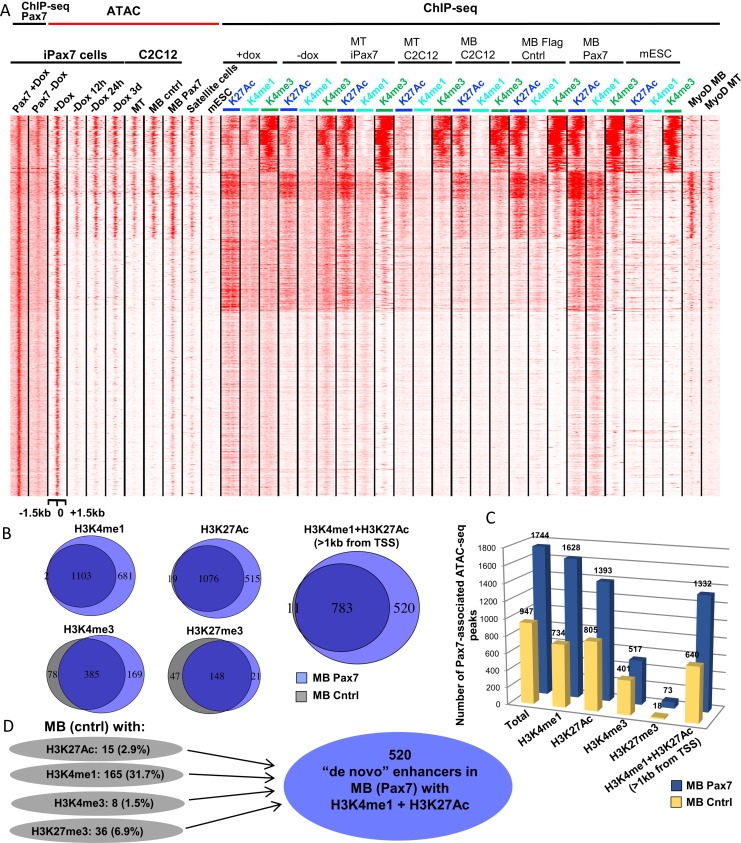
Pluripotent stem cells (PSC) hold great promise for the treatment of human skeletal muscle diseases. However, it remains challenging to convert PSC to skeletal muscle cells, and the mechanisms by which the master regulatory transcription factor, Pax7, promotes muscle stem (satellite) cell identity are not yet understood. We have taken advantage of PSC-derived skeletal muscle precursor cells (iPax7), wherein the induced expression of Pax7 robustly initiates the muscle program and enables the in vitro generation of precursors that seed the satellite cell compartment upon transplantation. Remarkably, we found that chromatin accessibility in myogenic precursors pre-figures subsequent activation of myogenic differentiation genes. We also found that Pax7 binding is generally restricted to euchromatic regions and excluded from H3K27 tri-methylated regions in muscle cells, suggesting that recruitment of this factor is circumscribed by chromatin state. Further, we show that Pax7 binding induces dramatic, localized remodeling of chromatin characterized by the acquisition of histone marks associated with enhancer activity and induction of chromatin accessibility in both muscle precursors and lineage-committed myoblasts. Conversely, removal of Pax7 leads to rapid reversal of these features on a subset of enhancers. Interestingly, another cluster of Pax7 binding sites is associated with a durably accessible and remodeled chromatin state after removal of Pax7, and persistent enhancer accessibility is associated with subsequent, proximal binding by the muscle regulatory factors, MyoD1 and myogenin. Our studies provide new insights into the epigenetic landscape of skeletal muscle stem cells and precursors and the role of Pax7 in satellite cell specification.
Conflict of interest statement
Figures

Similar articles
-
Alveolar rhabdomyosarcoma-associated proteins PAX3/FOXO1A and PAX7/FOXO1A suppress the transcriptional activity of MyoD-target genes in muscle stem cells.Oncogene. 2013 Jan 31;32(5):651-62. doi: 10.1038/onc.2012.73. Epub 2012 Jun 18. Oncogene. 2013. PMID: 22710712
-
Concise Review: Epigenetic Regulation of Myogenesis in Health and Disease.Stem Cells Transl Med. 2016 Mar;5(3):282-90. doi: 10.5966/sctm.2015-0266. Epub 2016 Jan 21. Stem Cells Transl Med. 2016. PMID: 26798058 Free PMC article. Review.
-
NEDD4 Regulates PAX7 Levels Promoting Activation of the Differentiation Program in Skeletal Muscle Precursors.Stem Cells. 2015 Oct;33(10):3138-51. doi: 10.1002/stem.2125. Epub 2015 Aug 25. Stem Cells. 2015. PMID: 26304770 Free PMC article.
-
mTOR is necessary for proper satellite cell activity and skeletal muscle regeneration.Biochem Biophys Res Commun. 2015 Jul 17-24;463(1-2):102-8. doi: 10.1016/j.bbrc.2015.05.032. Epub 2015 May 18. Biochem Biophys Res Commun. 2015. PMID: 25998386 Free PMC article.
-
The molecular regulation of muscle stem cell function.Cold Spring Harb Symp Quant Biol. 2008;73:323-31. doi: 10.1101/sqb.2008.73.064. Epub 2009 Mar 27. Cold Spring Harb Symp Quant Biol. 2008. PMID: 19329572 Review.
Cited by
-
The E3 ubiquitin ligase Nedd4L preserves skeletal muscle stem cell quiescence by inhibiting their activation.iScience. 2024 Jun 11;27(7):110241. doi: 10.1016/j.isci.2024.110241. eCollection 2024 Jul 19. iScience. 2024. PMID: 39015146 Free PMC article.
-
Skeletal Muscle Pathogenesis in Polyglutamine Diseases.Cells. 2022 Jul 3;11(13):2105. doi: 10.3390/cells11132105. Cells. 2022. PMID: 35805189 Free PMC article. Review.
-
Time-dependent Pax3-mediated chromatin remodeling and cooperation with Six4 and Tead2 specify the skeletal myogenic lineage in developing mesoderm.PLoS Biol. 2019 Feb 26;17(2):e3000153. doi: 10.1371/journal.pbio.3000153. eCollection 2019 Feb. PLoS Biol. 2019. PMID: 30807574 Free PMC article.
-
Muscle progenitor specification and myogenic differentiation are associated with changes in chromatin topology.Nat Commun. 2020 Dec 4;11(1):6222. doi: 10.1038/s41467-020-19999-w. Nat Commun. 2020. PMID: 33277476 Free PMC article.
-
Shaping Gene Expression by Landscaping Chromatin Architecture: Lessons from a Master.Mol Cell. 2018 Aug 2;71(3):375-388. doi: 10.1016/j.molcel.2018.04.025. Epub 2018 Jun 7. Mol Cell. 2018. PMID: 29887393 Free PMC article. Review.
References
-
- Darabi R, Gehlbach K, Bachoo RM, Kamath S, Osawa M, Kamm KE, et al. Functional skeletal muscle regeneration from differentiating embryonic stem cells. Nature medicine. 2008;14(2):134–43. doi: 10.1038/nm1705 - DOI - PubMed
-
- Hosoyama T, McGivern JV, Van Dyke JM, Ebert AD, Suzuki M. Derivation of myogenic progenitors directly from human pluripotent stem cells using a sphere-based culture. Stem cells translational medicine. 2014;3(5):564–74. doi: 10.5966/sctm.2013-0143 - DOI - PMC - PubMed
-
- Shelton M, Metz J, Liu J, Carpenedo RL, Demers SP, Stanford WL, et al. Derivation and expansion of PAX7-positive muscle progenitors from human and mouse embryonic stem cells. Stem cell reports. 2014;3(3):516–29. doi: 10.1016/j.stemcr.2014.07.001 - DOI - PMC - PubMed
-
- Xu C, Tabebordbar M, Iovino S, Ciarlo C, Liu J, Castiglioni A, et al. A zebrafish embryo culture system defines factors that promote vertebrate myogenesis across species. Cell. 2013;155(4):909–21. doi: 10.1016/j.cell.2013.10.023 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
