

The Cytoskeleton-Autophagy Connection
- PMID: 28441569
- PMCID: PMC5444402
- DOI: 10.1016/j.cub.2017.02.061
The Cytoskeleton-Autophagy Connection
Abstract
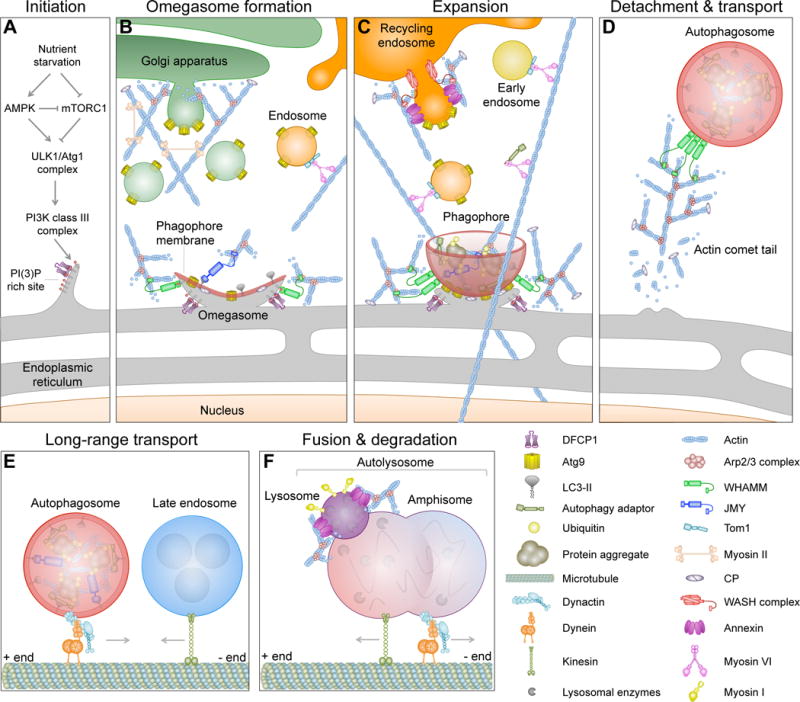
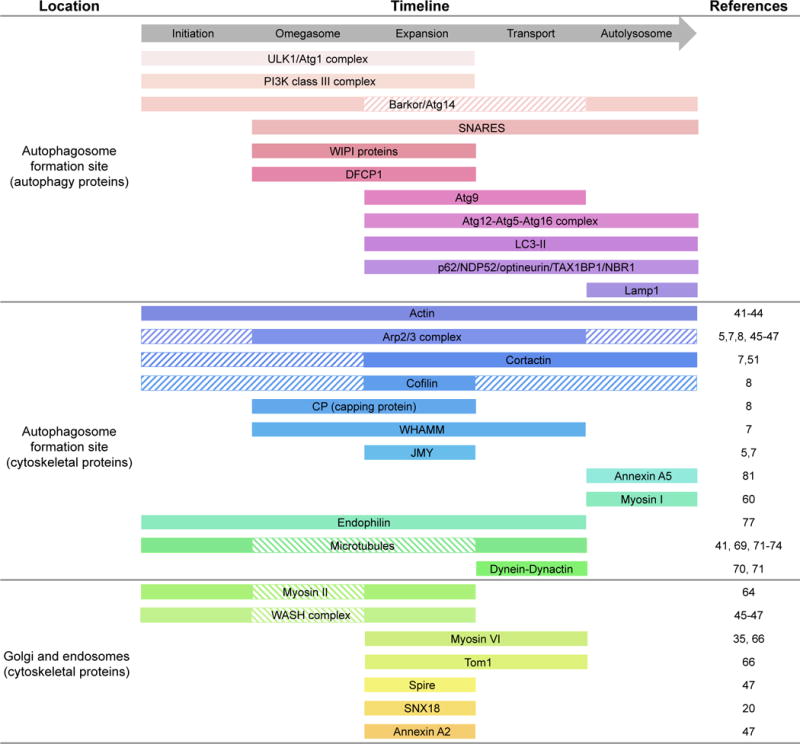
Actin cytoskeleton dynamics play vital roles in most forms of intracellular trafficking by promoting the biogenesis and transport of vesicular cargoes. Mounting evidence indicates that actin dynamics and membrane-cytoskeleton scaffolds also have essential roles in macroautophagy, the process by which cellular waste is isolated inside specialized vesicles called autophagosomes for recycling and degradation. Branched actin polymerization is necessary for the biogenesis of autophagosomes from the endoplasmic reticulum (ER) membrane. Actomyosin-based transport is then used to feed the growing phagophore with pre-selected cargoes and debris derived from different membranous organelles inside the cell. Finally, mature autophagosomes detach from the ER membrane by an as yet unknown mechanism, undergo intracellular transport and then fuse with lysosomes, endosomes and multivesicular bodies through mechanisms that involve actin- and microtubule-mediated motility, cytoskeleton-membrane scaffolds and signaling proteins. In this review, we highlight the considerable progress made recently towards understanding the diverse roles of the cytoskeleton in autophagy.
Published by Elsevier Ltd.
Figures
References
-
- Anitei M, Hoflack B. Bridging membrane and cytoskeleton dynamics in the secretory and endocytic pathways. Nat Cell Biol. 2011;14:11–19. - PubMed
-
- Suetsugu S, Gautreau A. Synergistic BAR-NPF interactions in actin-driven membrane remodeling. Trends Cell Biol. 2012;22:141–150. - PubMed
-
- Kaksonen M, Toret CP, Drubin DG. Harnessing actin dynamics for clathrin-mediated endocytosis. Nat Rev Mol Cell Biol. 2006;7:404–414. - PubMed
-
- Egea G, Lazaro-Dieguez F, Vilella M. Actin dynamics at the Golgi complex in mammalian cells. Curr Opin Cell Biol. 2006;18:168–178. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
