

Chlamydia trachomatis neither exerts deleterious effects on spermatozoa nor impairs male fertility
- PMID: 28442719
- PMCID: PMC5430866
- DOI: 10.1038/s41598-017-01262-w
Chlamydia trachomatis neither exerts deleterious effects on spermatozoa nor impairs male fertility
Erratum in
-
Erratum: Chlamydia trachomatis neither exerts deleterious effects on spermatozoa nor impairs male fertility.Sci Rep. 2017 Nov 21;7(1):16312. doi: 10.1038/s41598-017-12316-4. Sci Rep. 2017. PMID: 29162849 Free PMC article.
Abstract
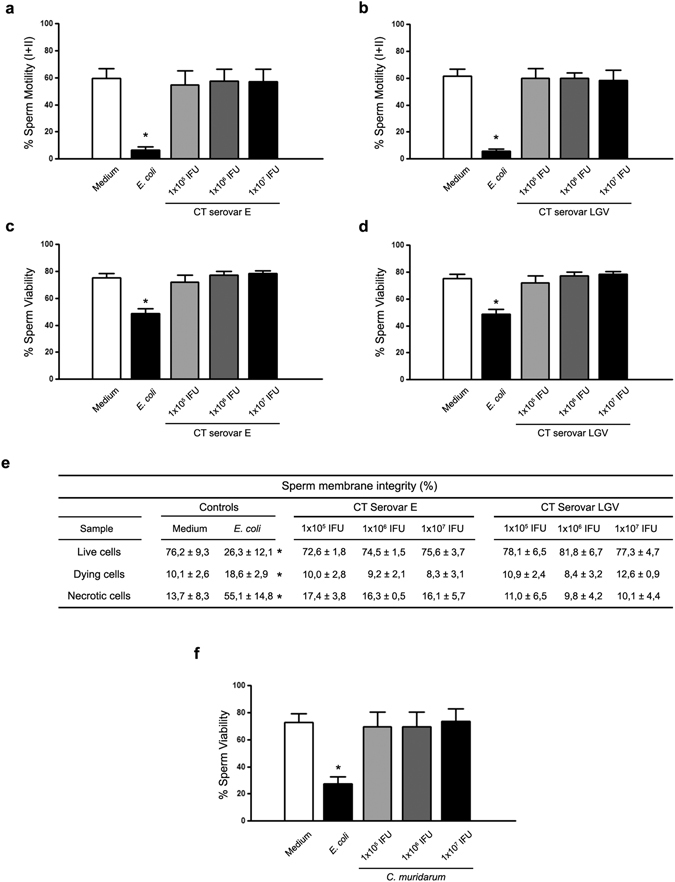
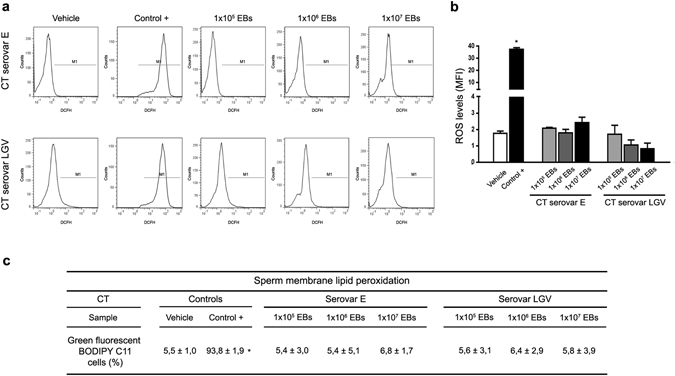
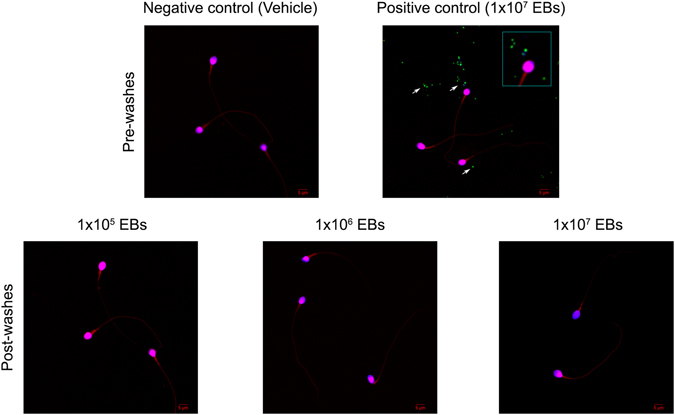
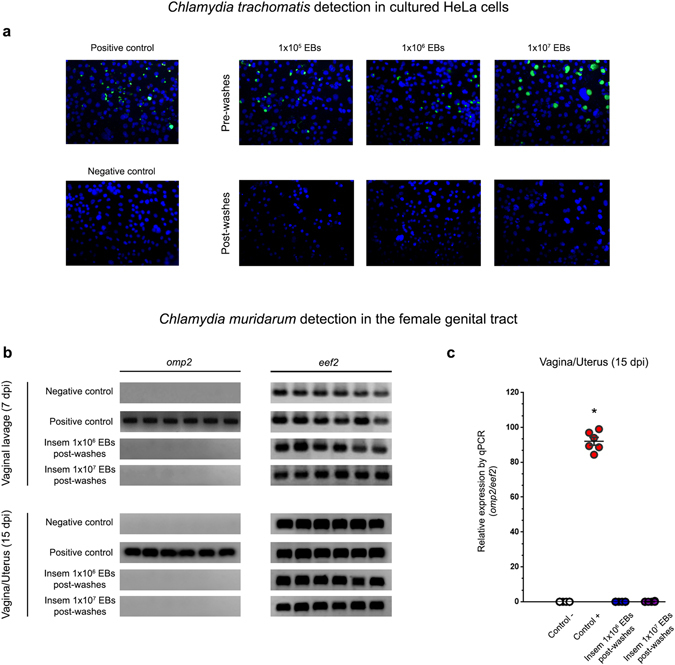
Chlamydia trachomatis is the most prevalent sexually transmitted bacterial infection. However, whether Chlamydia trachomatis has a negative impact on sperm quality and male fertility is still controversial. Herein, we report the effects on sperm quality of the in vitro exposure of spermatozoa to Chlamydia trachomatis, and also the effects of male genital infection on male fertility using an animal model. Human and mouse sperm were obtained from healthy donors and cauda epididimys from C57BL/6 mice, respectively. Highly motile human or mouse spermatozoa were in vitro exposed to C. trachomatis (serovar E or LGV) or C. muridarum, respectively. Then, sperm quality parameters were analyzed. Moreover, male fertility of Chlamydia muridarum infected male C57BL/6 mice was assessed. Human or murine sperm in vitro exposed to increasing bacterial concentrations or soluble factors from C. trachomatis or C. muridarum, respectively, did not show differences in sperm motility and viability, apoptosis, mitochondrial membrane potential, DNA fragmentation, ROS production and lipid peroxidation levels, when compared with control sperm (p > 0.05). Moreover, no differences in fertility parameters (potency, fecundity, fertility index, pre- and post-implantation loss) were observed between control and infected males. In conclusion, our results indicate that Chlamydia spp. neither directly exerts deleterious effects on spermatozoa nor impairs male fertility.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
