

Phosphorylation of the mitochondrial autophagy receptor Nix enhances its interaction with LC3 proteins
- PMID: 28442745
- PMCID: PMC5430633
- DOI: 10.1038/s41598-017-01258-6
Phosphorylation of the mitochondrial autophagy receptor Nix enhances its interaction with LC3 proteins
Abstract
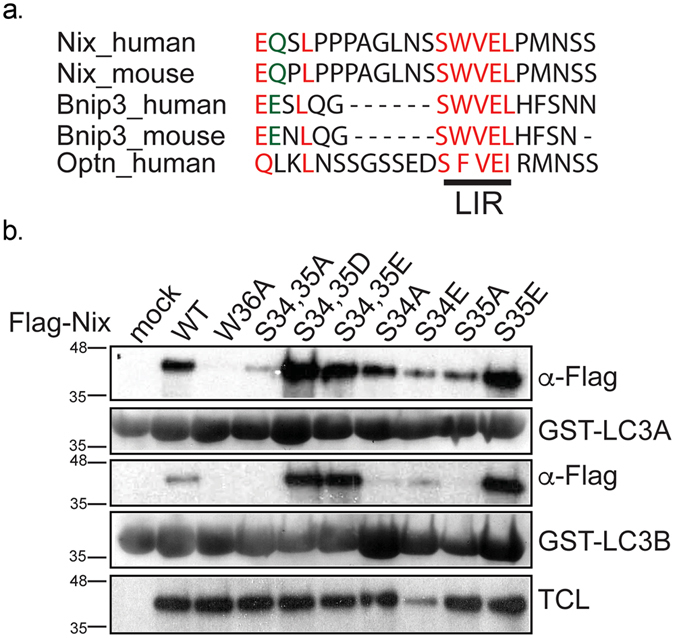
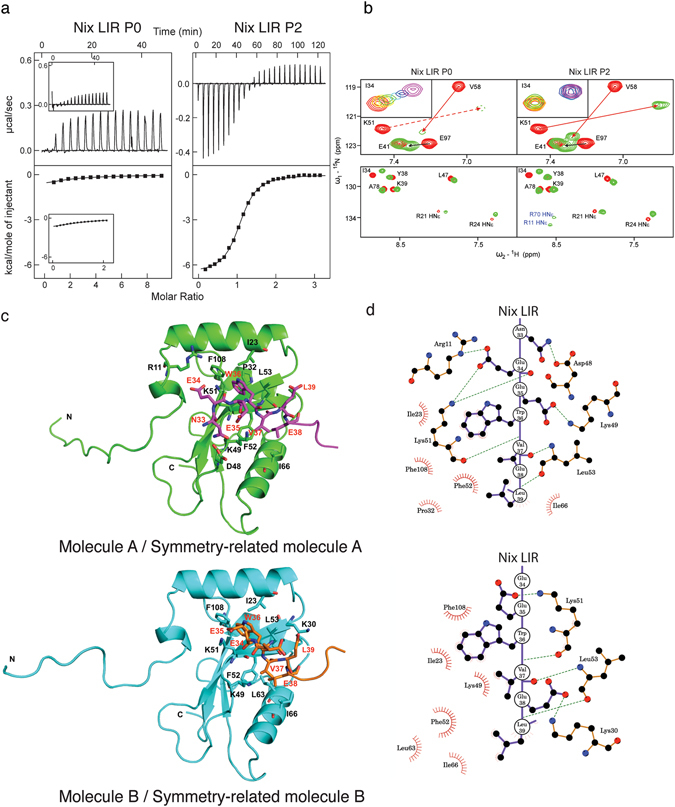
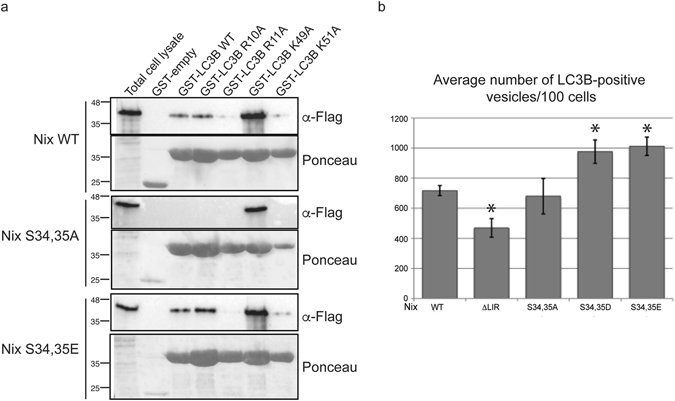
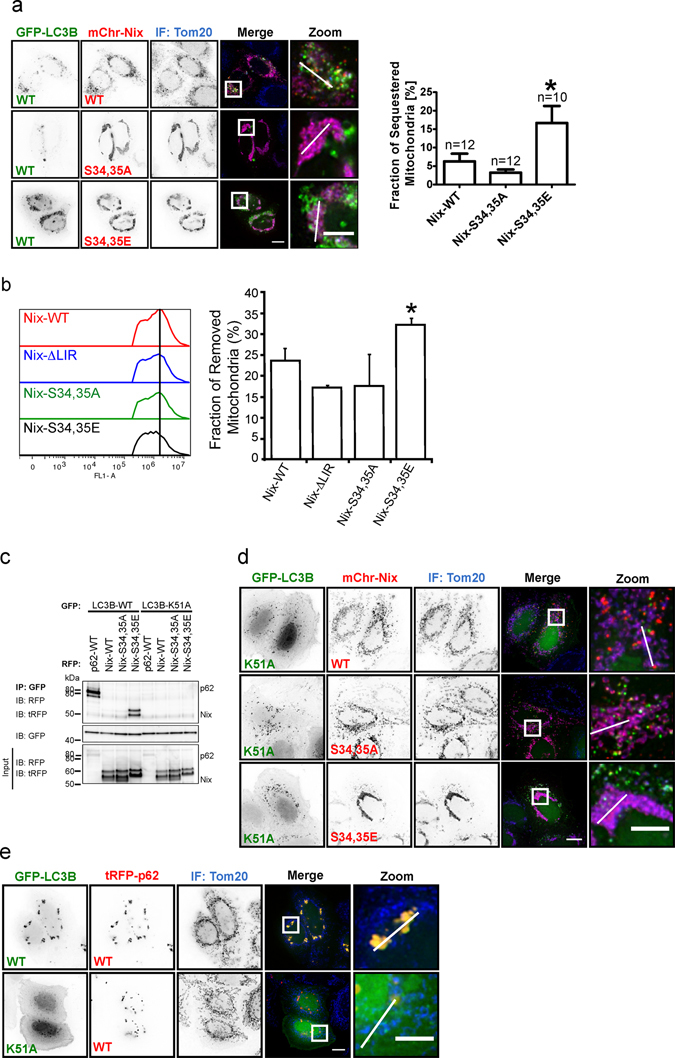
The mitophagy receptor Nix interacts with LC3/GABARAP proteins, targeting mitochondria into autophagosomes for degradation. Here we present evidence for phosphorylation-driven regulation of the Nix:LC3B interaction. Isothermal titration calorimetry and NMR indicate a ~100 fold enhanced affinity of the serine 34/35-phosphorylated Nix LC3-interacting region (LIR) to LC3B and formation of a very rigid complex compared to the non-phosphorylated sequence. Moreover, the crystal structure of LC3B in complex with the Nix LIR peptide containing glutamic acids as phosphomimetic residues and NMR experiments revealed that LIR phosphorylation stabilizes the Nix:LC3B complex via formation of two additional hydrogen bonds between phosphorylated serines of Nix LIR and Arg11, Lys49 and Lys51 in LC3B. Substitution of Lys51 to Ala in LC3B abrogates binding of a phosphomimetic Nix mutant. Functionally, serine 34/35 phosphorylation enhances autophagosome recruitment to mitochondria in HeLa cells. Together, this study provides cellular, biochemical and biophysical evidence that phosphorylation of the LIR domain of Nix enhances mitophagy receptor engagement.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
