

Interplay between recombinant Hsp70 and proteasomes: proteasome activity modulation and ubiquitin-independent cleavage of Hsp70
- PMID: 28447215
- PMCID: PMC5573687
- DOI: 10.1007/s12192-017-0792-y
Interplay between recombinant Hsp70 and proteasomes: proteasome activity modulation and ubiquitin-independent cleavage of Hsp70
Abstract
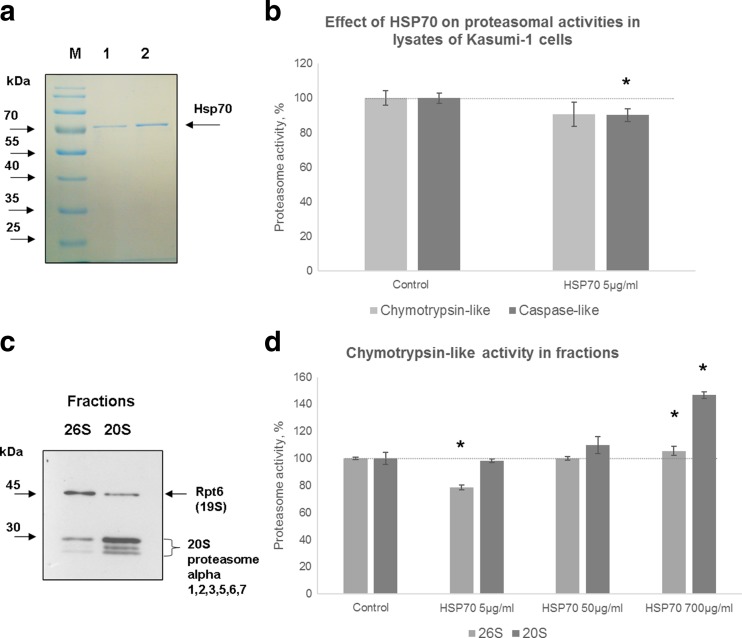
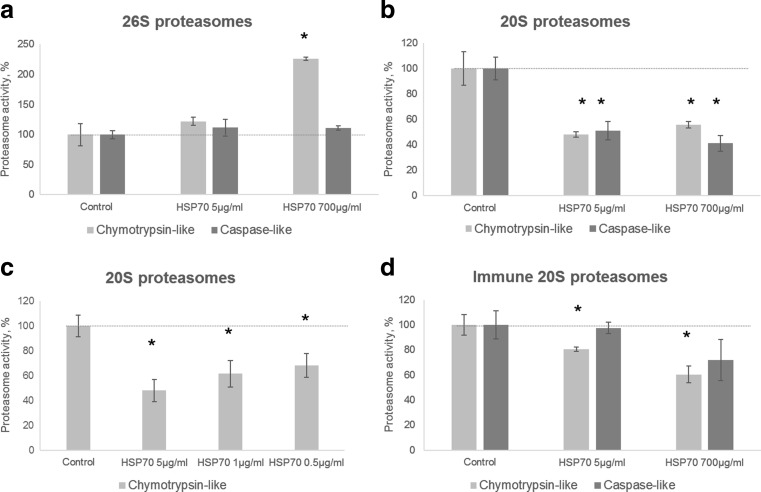
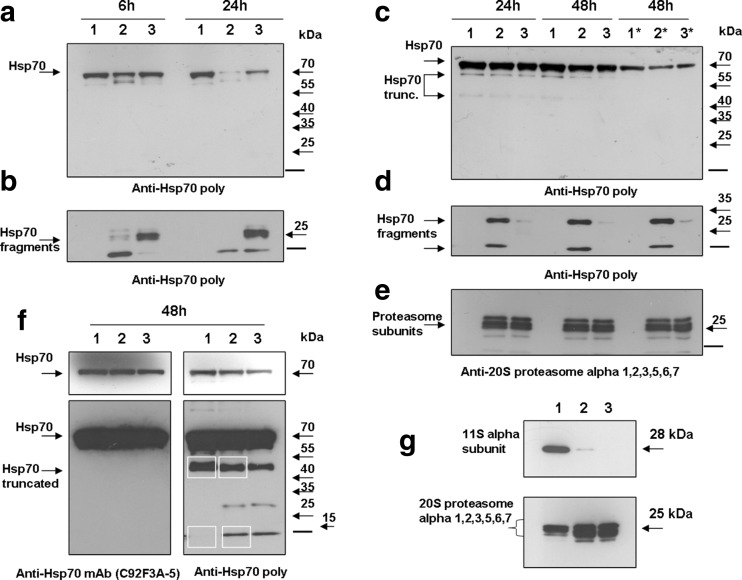
The heat shock protein 70 (Hsp70, human HSPA1A) plays indispensable roles in cellular stress responses and protein quality control (PQC). In the framework of PQC, it cooperates with the ubiquitin-proteasome system (UPS) to clear damaged and dysfunctional proteins in the cell. Moreover, Hsp70 itself is rapidly degraded following the recovery from stress. It was demonstrated that its fast turnover is mediated via ubiquitination and subsequent degradation by the 26S proteasome. At the same time, the effect of Hsp70 on the functional state of proteasomes has been insufficiently investigated. Here, we characterized the direct effect of recombinant Hsp70 on the activity of 20S and 26S proteasomes and studied Hsp70 degradation by the 20S proteasome in vitro. We have shown that the activity of purified 20S proteasomes is decreased following incubation with recombinant human Hsp70. On the other hand, high concentrations of Hsp70 activated 26S proteasomes. Finally, we obtained evidence that in addition to previously reported ubiquitin-dependent degradation, Hsp70 could be cleaved independent of ubiquitination by the 20S proteasome. The results obtained reveal novel aspects of the interplay between Hsp70 and proteasomes.
Keywords: Heat shock proteins; Hsp70; Proteasome; Proteasome regulators; Ubiquitin-independent degradation.
Conflict of interest statement
The investigation of proteasome activity in the culture cells and extracts was supported by Ministry of Education and Science of Russian Federation (agreement no. 14.Z50.31.0014) Russian Science Foundation grant (14-50-00060) and Russian President Foundation grant (МК-3613.2017.4) to A.M.
Figures
References
-
- Abramova EB, Astakhova TM, Erokhov PA, Sharova NP (2004) Multiple forms of the proteasomes and some approaches to their separation Izvestiia Akademii nauk Seriia biologicheskaia / Rossiiskaia akademiia nauk:150–156 - PubMed
-
- Abramova EB, Sharova NP, Karpov VL. The proteasome: destroy to live. Mol Biol. 2002;36:761–776. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
