

IL-33/ST2 pathway drives regulatory T cell dependent suppression of liver damage upon cytomegalovirus infection
- PMID: 28448566
- PMCID: PMC5423658
- DOI: 10.1371/journal.ppat.1006345
IL-33/ST2 pathway drives regulatory T cell dependent suppression of liver damage upon cytomegalovirus infection
Abstract
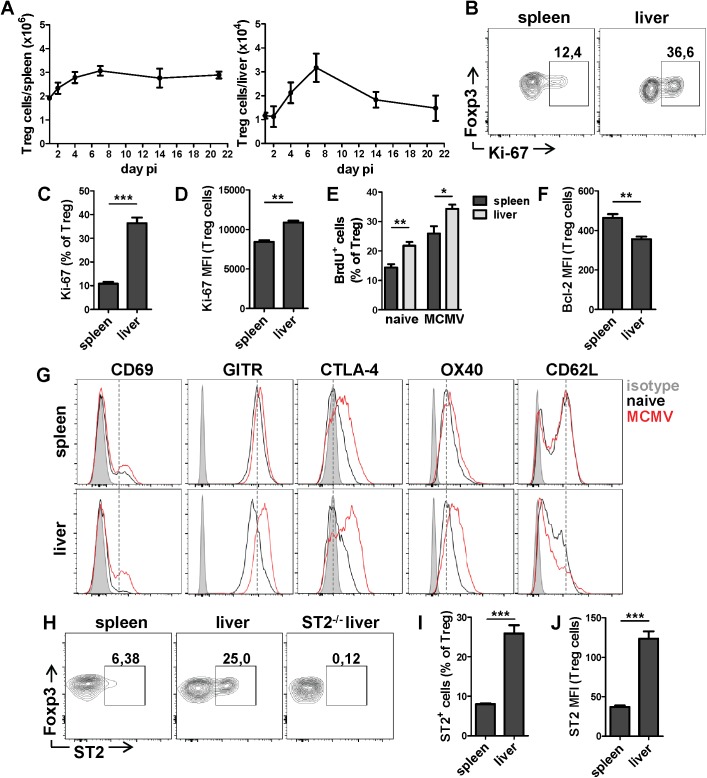
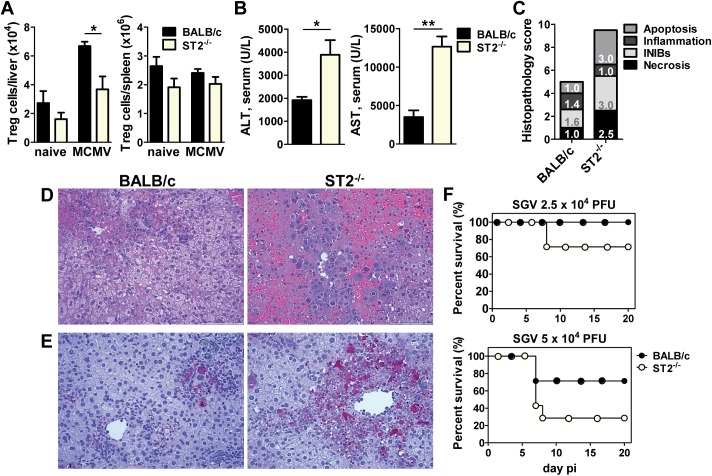
Regulatory T (Treg) cells dampen an exaggerated immune response to viral infections in order to avoid immunopathology. Cytomegaloviruses (CMVs) are herpesviruses usually causing asymptomatic infection in immunocompetent hosts and induce strong cellular immunity which provides protection against CMV disease. It remains unclear how these persistent viruses manage to avoid induction of immunopathology not only during the acute infection but also during life-long persistence and virus reactivation. This may be due to numerous viral immunoevasion strategies used to specifically modulate immune responses but also induction of Treg cells by CMV infection. Here we demonstrate that liver Treg cells are strongly induced in mice infected with murine CMV (MCMV). The depletion of Treg cells results in severe hepatitis and liver damage without alterations in the virus load. Moreover, liver Treg cells show a high expression of ST2, a cellular receptor for tissue alarmin IL-33, which is strongly upregulated in the liver of infected mice. We demonstrated that IL-33 signaling is crucial for Treg cell accumulation after MCMV infection and ST2-deficient mice show a more pronounced liver pathology and higher mortality compared to infected control mice. These results illustrate the importance of IL-33 in the suppressive function of liver Treg cells during CMV infection.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures







References
-
- Josefowicz SZ, Lu LF, Rudensky AY. Regulatory T cells: mechanisms of differentiation and function. Annu Rev Immunol. 2012;30:531–64. doi: 10.1146/annurev.immunol.25.022106.141623 - DOI - PMC - PubMed
-
- Curotto de Lafaille MA, Lafaille JJ. Natural and adaptive foxp3+ regulatory T cells: more of the same or a division of labor? Immunity. 2009;30(5):626–35. doi: 10.1016/j.immuni.2009.05.002 - DOI - PubMed
-
- Cretney E, Kallies A, Nutt SL. Differentiation and function of Foxp3(+) effector regulatory T cells. Trends Immunol. 2013;34(2):74–80. doi: 10.1016/j.it.2012.11.002 - DOI - PubMed
-
- Burzyn D, Benoist C, Mathis D. Regulatory T cells in nonlymphoid tissues. Nat Immunol. 2013;14(10):1007–13. PubMed Central PMCID: PMCPMC4708287. doi: 10.1038/ni.2683 - DOI - PMC - PubMed
-
- Arpaia N, Green JA, Moltedo B, Arvey A, Hemmers S, Yuan S, et al. A Distinct Function of Regulatory T Cells in Tissue Protection. Cell. 2015;162(5):1078–89. PubMed Central PMCID: PMCPMC4603556. doi: 10.1016/j.cell.2015.08.021 - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
