

The Interplay of Cofactor Interactions and Post-translational Modifications in the Regulation of the AAA+ ATPase p97
- PMID: 28451587
- PMCID: PMC5389986
- DOI: 10.3389/fmolb.2017.00021
The Interplay of Cofactor Interactions and Post-translational Modifications in the Regulation of the AAA+ ATPase p97
Abstract
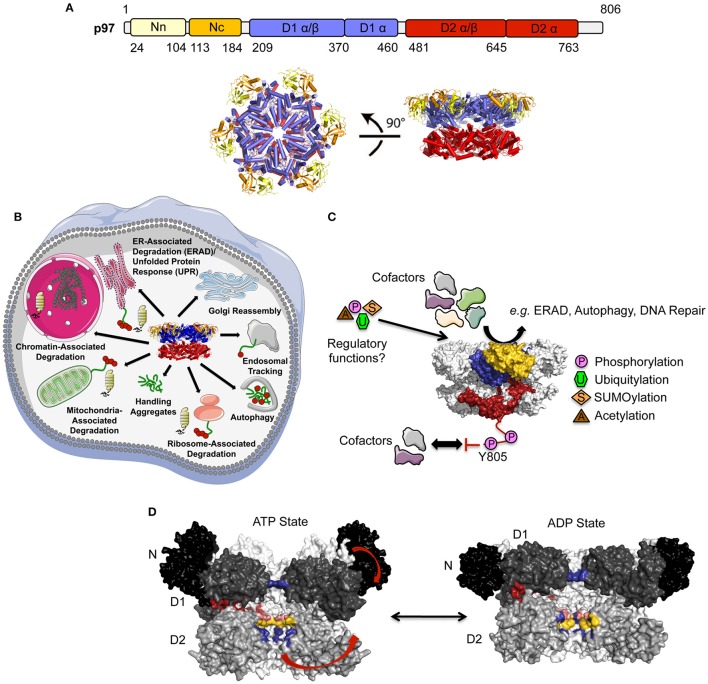
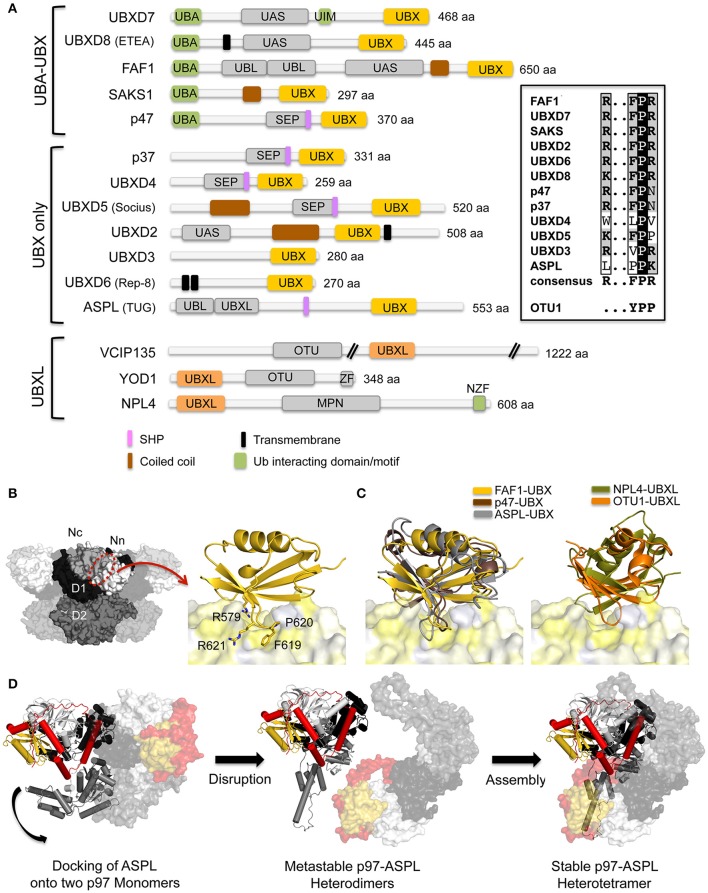
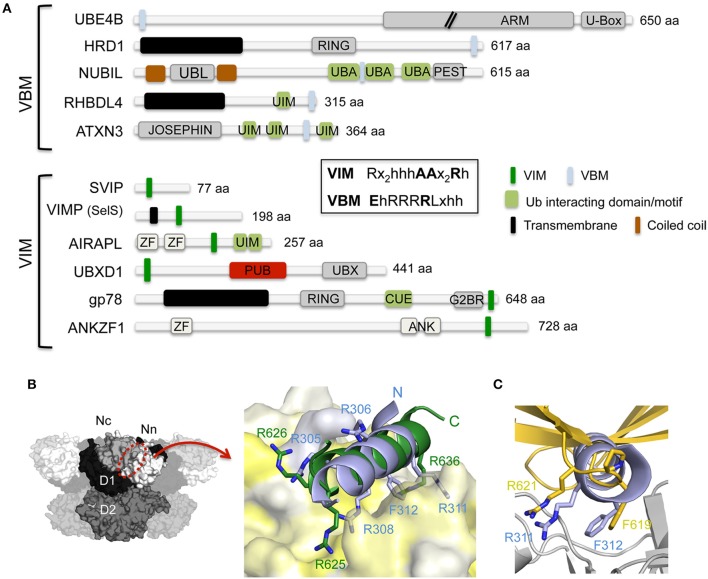
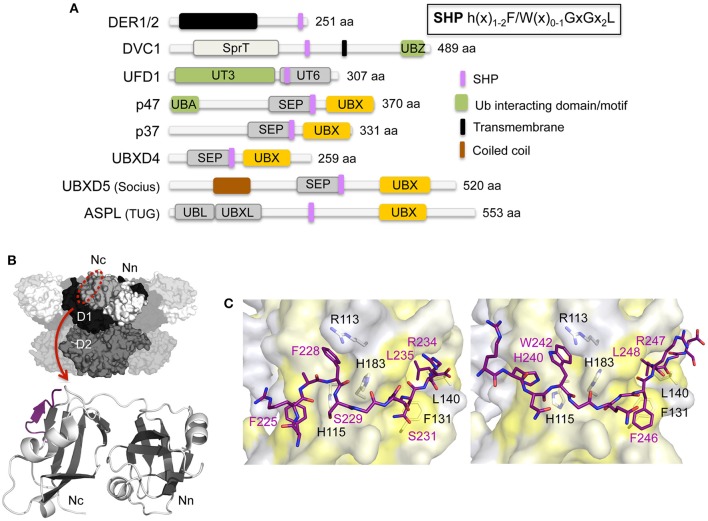
The hexameric type II AAA ATPase (ATPase associated with various activities) p97 (also referred to as VCP, Cdc48, and Ter94) is critically involved in a variety of cellular activities including pathways such as DNA replication and repair which both involve chromatin remodeling, and is a key player in various protein quality control pathways mediated by the ubiquitin proteasome system as well as autophagy. Correspondingly, p97 has been linked to various pathophysiological states including cancer, neurodegeneration, and premature aging. p97 encompasses an N-terminal domain, two highly conserved ATPase domains and an unstructured C-terminal tail. This enzyme hydrolyzes ATP and utilizes the resulting energy to extract or disassemble protein targets modified with ubiquitin from stable protein assemblies, chromatin and membranes. p97 participates in highly diverse cellular processes and hence its activity is tightly controlled. This is achieved by multiple regulatory cofactors, which either associate with the N-terminal domain or interact with the extreme C-terminus via distinct binding elements and target p97 to specific cellular pathways, sometimes requiring the simultaneous association with more than one cofactor. Most cofactors are recruited to p97 through conserved binding motifs/domains and assist in substrate recognition or processing by providing additional molecular properties. A tight control of p97 cofactor specificity and diversity as well as the assembly of higher-order p97-cofactor complexes is accomplished by various regulatory mechanisms, which include bipartite binding, binding site competition, changes in oligomeric assemblies, and nucleotide-induced conformational changes. Furthermore, post-translational modifications (PTMs) like acetylation, palmitoylation, phosphorylation, SUMOylation, and ubiquitylation of p97 have been reported which further modulate its diverse molecular activities. In this review, we will describe the molecular basis of p97-cofactor specificity/diversity and will discuss how PTMs can modulate p97-cofactor interactions and affect the physiological and patho-physiological functions of p97.
Keywords: AAA+ ATPase; cofactor diversity; conformational changes; p97; post-translational modification; protein disassembly; protein quality control.
Figures
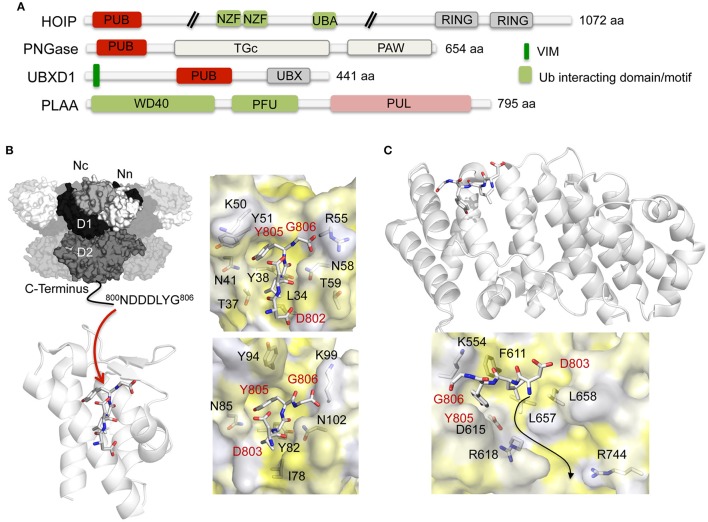
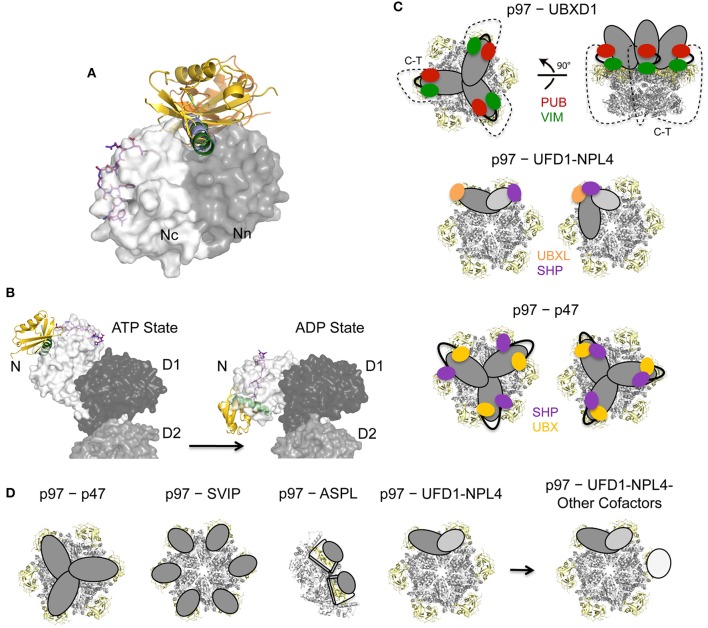

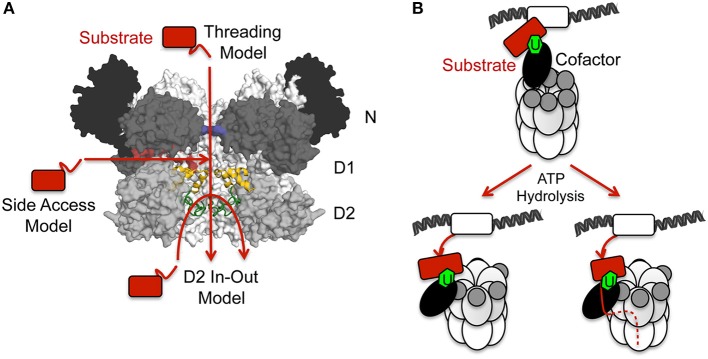
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous