

Daily Light Exposure Patterns Reveal Phase and Period of the Human Circadian Clock
- PMID: 28452285
- PMCID: PMC5476188
- DOI: 10.1177/0748730417696787
Daily Light Exposure Patterns Reveal Phase and Period of the Human Circadian Clock
Abstract
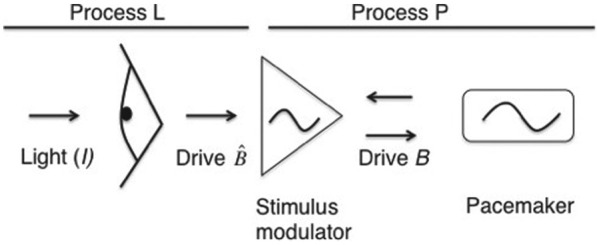
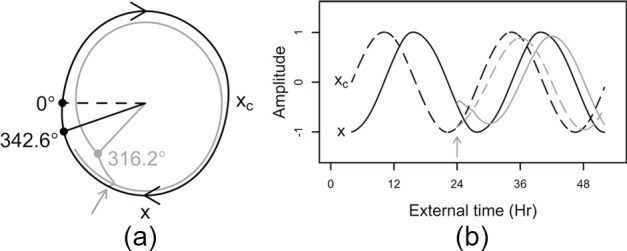
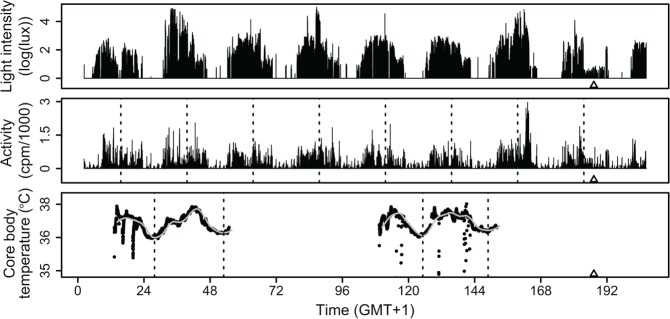
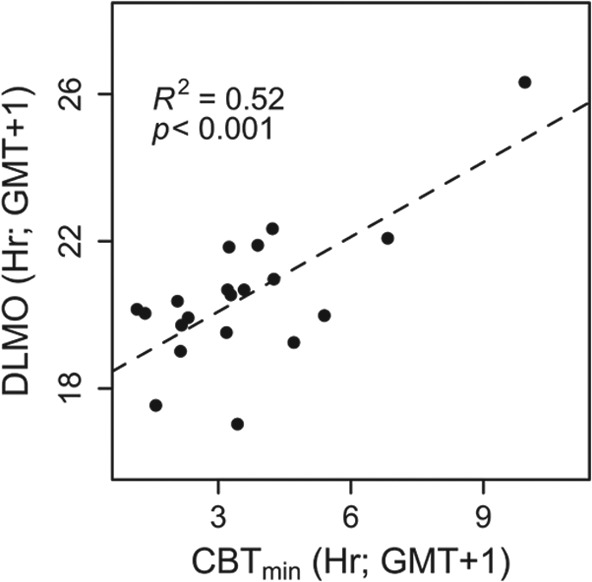
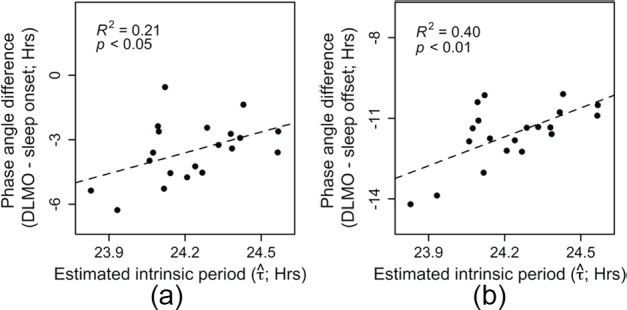
Light is the most potent time cue that synchronizes (entrains) the circadian pacemaker to the 24-h solar cycle. This entrainment process is an interplay between an individual's daily light perception and intrinsic pacemaker period under free-running conditions. Establishing individual estimates of circadian phase and period can be time-consuming. We show that circadian phase can be accurately predicted (SD = 1.1 h for dim light melatonin onset, DLMO) using 9 days of ambulatory light and activity data as an input to Kronauer's limit-cycle model for the human circadian system. This approach also yields an estimated circadian period of 24.2 h (SD = 0.2 h), with longer periods resulting in later DLMOs. A larger amount of daylight exposure resulted in an earlier DLMO. Individuals with a long circadian period also showed shorter intervals between DLMO and sleep timing. When a field-based estimation of tau can be validated under laboratory studies in a wide variety of individuals, the proposed methods may prove to be essential tools for individualized chronotherapy and light treatment for shift work and jetlag applications. These methods may improve our understanding of fundamental properties of human circadian rhythms under daily living conditions.
Keywords: CBT; DLMO; activity; ambulatory; circadian pacemaker; core body temperature; intrinsic period; light; modeling; tau.
Conflict of interest statement
Figures
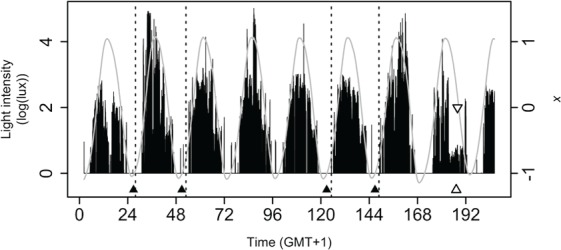
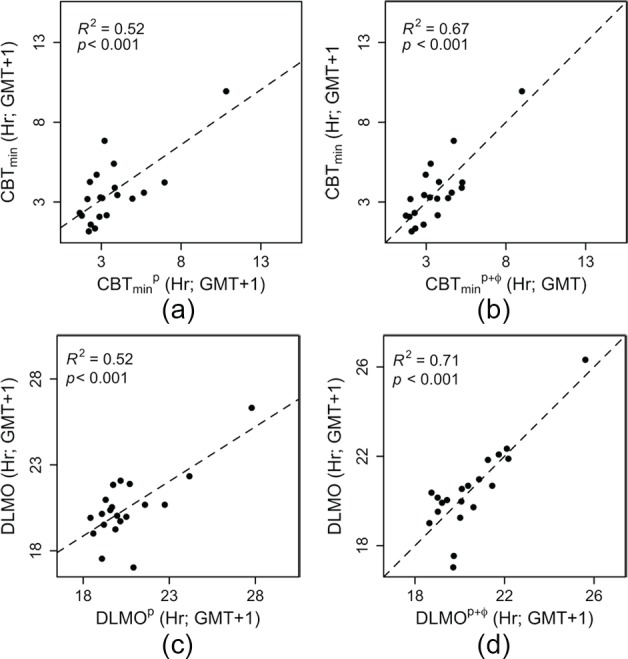
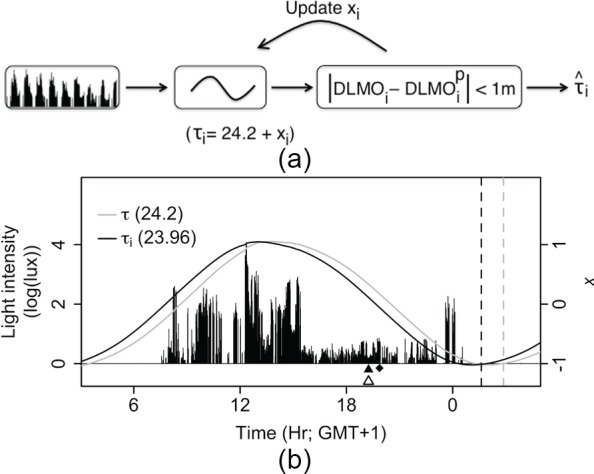
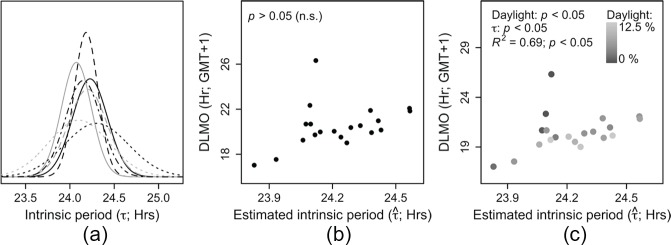
References
-
- Bates D, Mächler M, Bolker B. (2012). Fitting linear mixed-effects models using lme4. J Stat Softw 51:1-48.
-
- Beck AT, Steer RA, Brown GK. (1996). Manual for the Beck Depression Inventory—II. San Antonio, TX: Psychological Corporation.
-
- Benloucif S, Guico MJ, Reid KJ, Wolfe LF, L’hermite-Balériaux M, Zee PC. (2005). Stability of melatonin and temperature as circadian phase markers and their relation to sleep times in humans. J Biol Rhythms 20(2):178-188. - PubMed
-
- Brown EN, Choe Y, Shanahan TL, Czeisler CA. (1997). A mathematical model of diurnal variations in human plasma melatonin levels. Am J Physiol 272(3 Pt 1):E506-516. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
