

Acetylation of lysine ϵ-amino groups regulates aminoacyl-tRNA synthetase activity in Escherichia coli
- PMID: 28455447
- PMCID: PMC5481575
- DOI: 10.1074/jbc.M116.770826
Acetylation of lysine ϵ-amino groups regulates aminoacyl-tRNA synthetase activity in Escherichia coli
Abstract
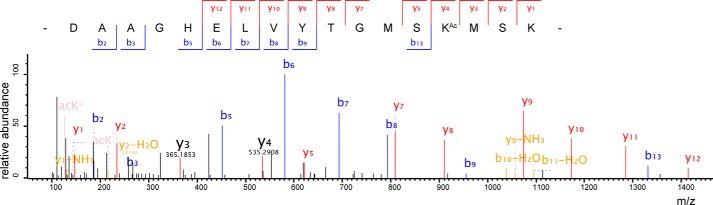
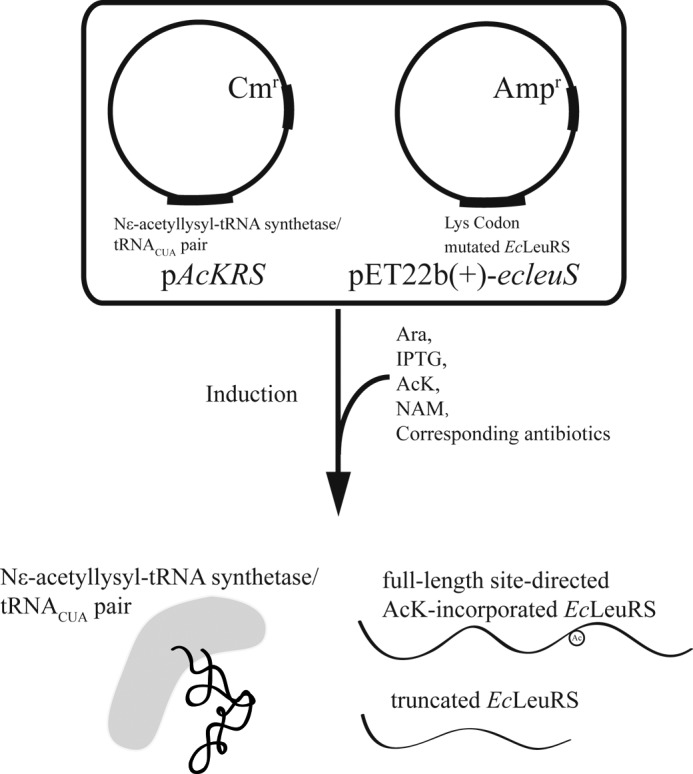
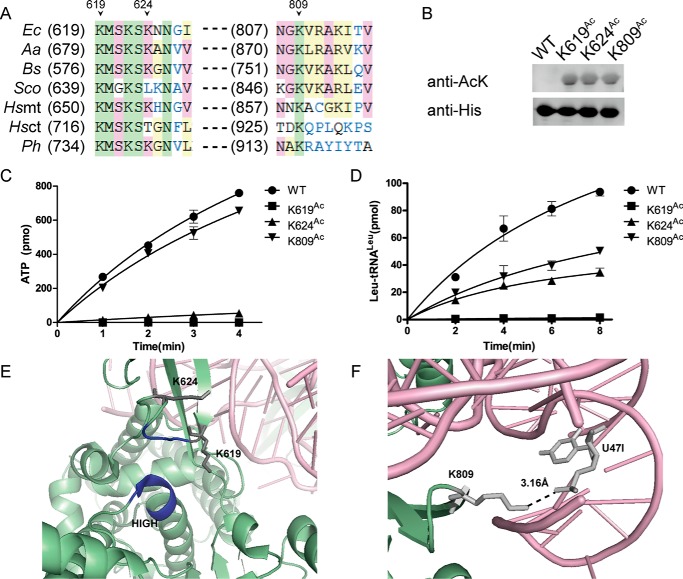
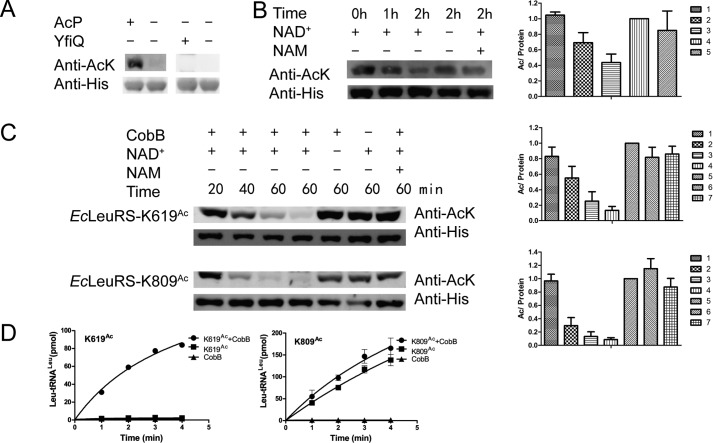
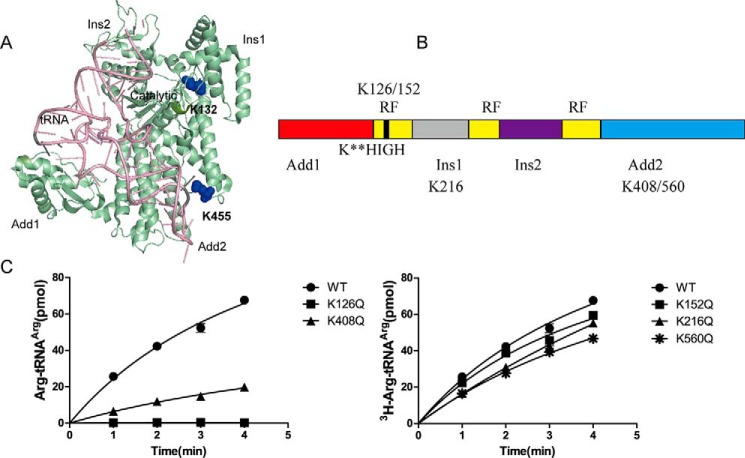
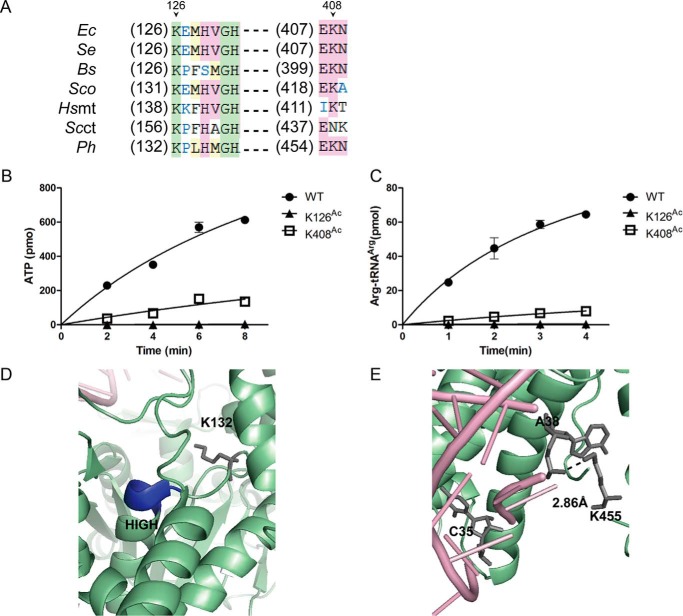
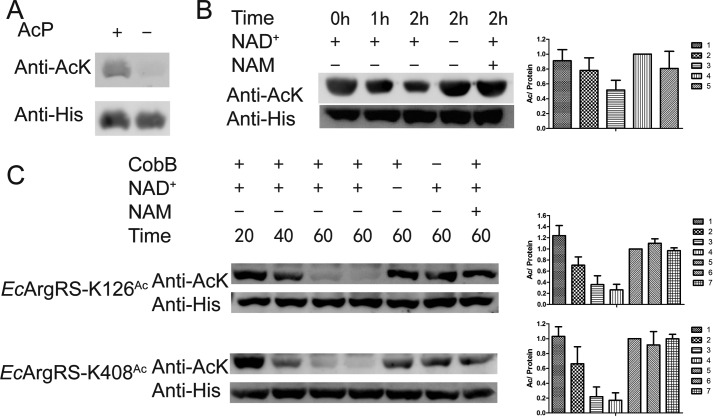
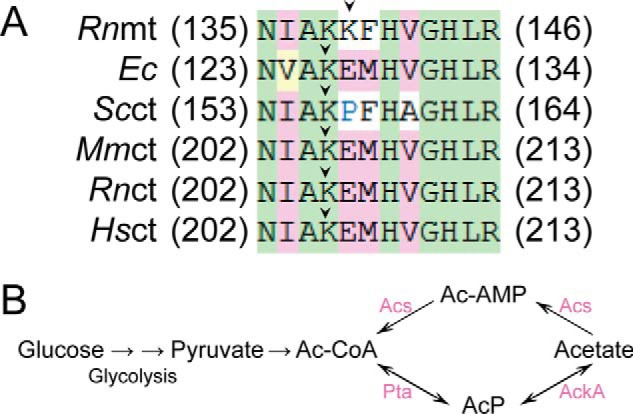
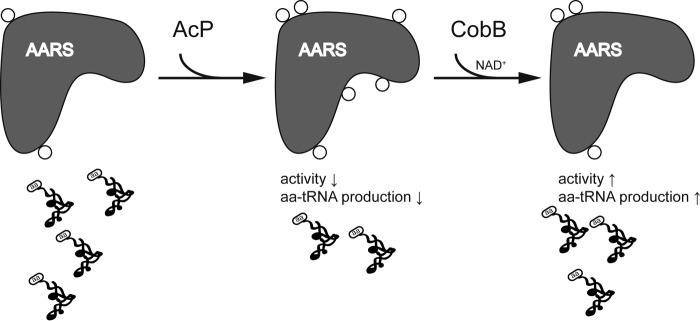
Previous proteomic analyses have shown that aminoacyl-tRNA synthetases in many organisms can be modified by acetylation of Lys. In this present study, leucyl-tRNA synthetase and arginyl-tRNA synthetase from Escherichia coli (EcLeuRS and EcArgRS) were overexpressed and purified and found to be acetylated on Lys residues by MS. Gln scanning mutagenesis revealed that Lys619, Lys624, and Lys809 in EcLeuRS and Lys126 and Lys408 in EcArgRS might play important roles in enzyme activity. Furthermore, we utilized a novel protein expression system to obtain enzymes harboring acetylated Lys at specific sites and investigated their catalytic activity. Acetylation of these Lys residues could affect their aminoacylation activity by influencing amino acid activation and/or the affinity for tRNA. In vitro assays showed that acetyl-phosphate nonenzymatically acetylates EcLeuRS and EcArgRS and suggested that the sirtuin class deacetylase CobB might regulate acetylation of these two enzymes. These findings imply a potential regulatory role for Lys acetylation in controlling the activity of aminoacyl-tRNA synthetases and thus protein synthesis.
Keywords: acetylation; amino acid; aminoacyl tRNA synthetase; catalysis; transfer RNA (tRNA); translation.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures


References
-
- Nangle L. A., De Crecy Lagard V., Doring V., and Schimmel P. (2002) Genetic code ambiguity: cell viability related to the severity of editing defects in mutant tRNA synthetases. J. Biol. Chem. 277, 45729–45733 - PubMed
-
- Lee J. W., Beebe K., Nangle L. A., Jang J., Longo-Guess C. M., Cook S. A., Davisson M. T., Sundberg J. P., Schimmel P., and Ackerman S. L. (2006) Editing-defective tRNA synthetase causes protein misfolding and neurodegeneration. Nature 443, 50–55 - PubMed
-
- Nangle L. A., Motta C. M., and Schimmel P. (2006) Global effects of mistranslation from an editing defect in mammalian cells. Chem. Biol. 13, 1091–1100 - PubMed
-
- Eriani G., Delarue M., Poch O., Gangloff J., and Moras D. (1990) Partition of tRNA synthetases into two classes based on mutually exclusive sets of sequence motifs. Nature 347, 203–206 - PubMed
-
- Ibba M., and Soll D. (2000) Aminoacyl-tRNA synthesis. Annu. Rev. Biochem. 69, 617–650 - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
