

Molecular impact of covalent modifications on nonribosomal peptide synthetase carrier protein communication
- PMID: 28455448
- PMCID: PMC5473208
- DOI: 10.1074/jbc.M116.766220
Molecular impact of covalent modifications on nonribosomal peptide synthetase carrier protein communication
Abstract
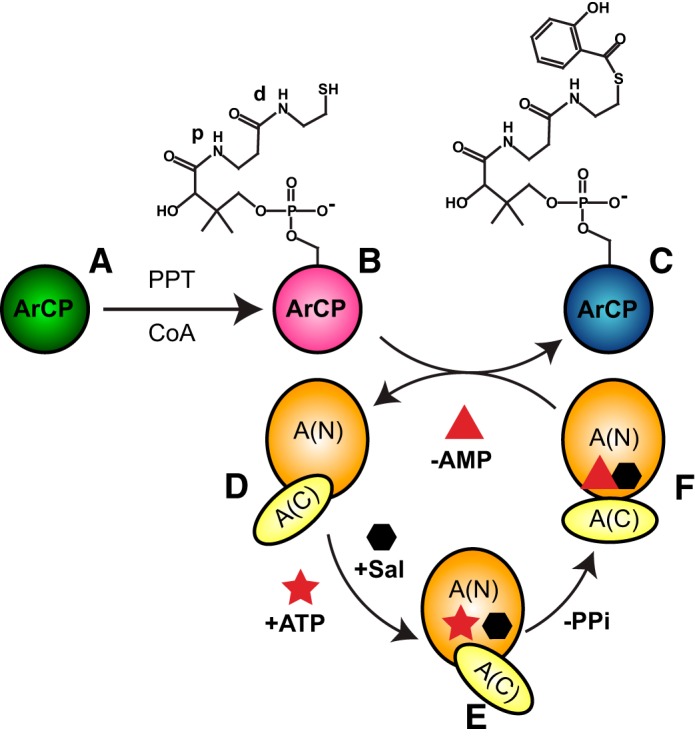
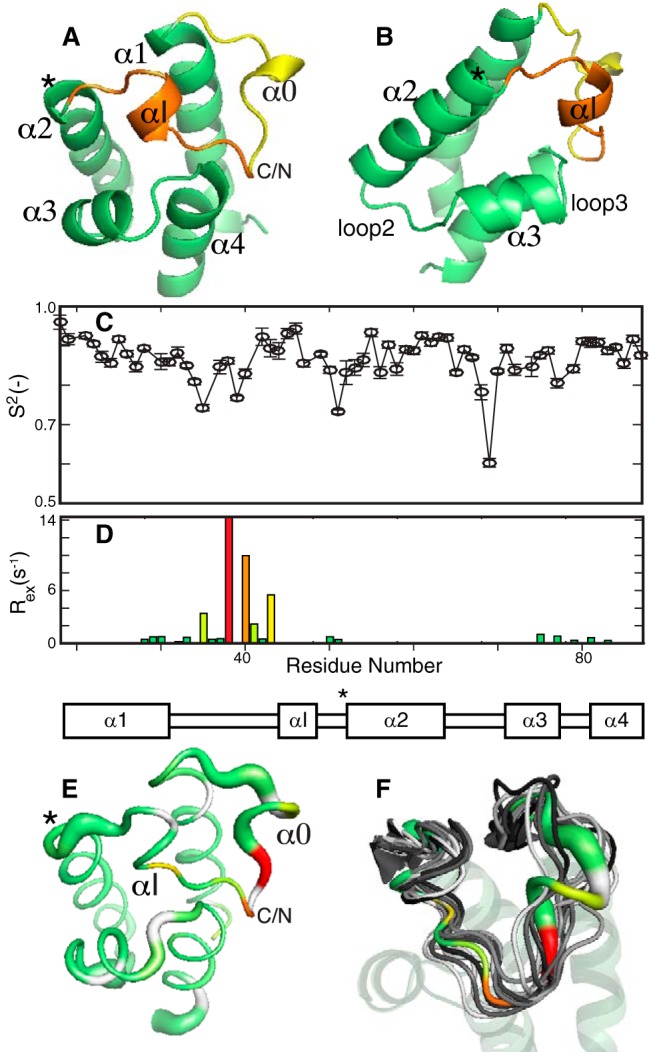
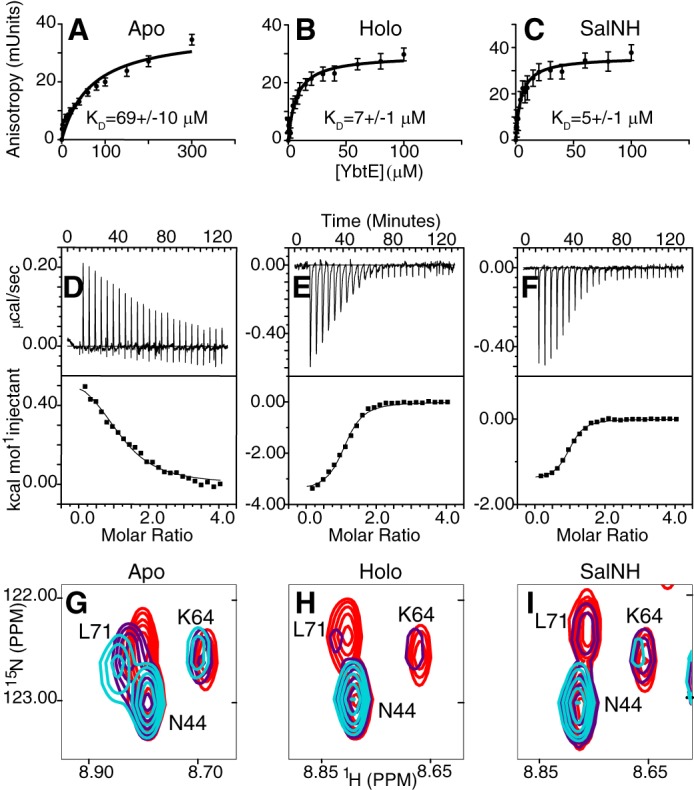
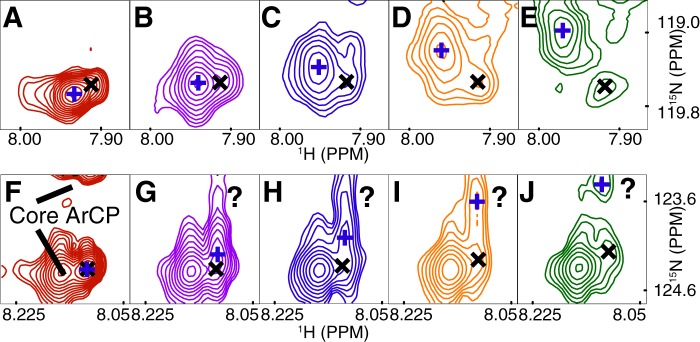
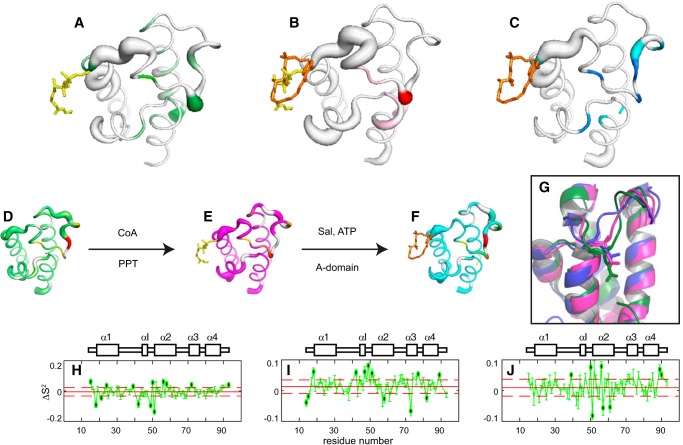
Nonribosomal peptide synthesis involves the interplay between covalent protein modifications, conformational fluctuations, catalysis, and transient protein-protein interactions. Delineating the mechanisms involved in orchestrating these various processes will deepen our understanding of domain-domain communication in nonribosomal peptide synthetases (NRPSs) and lay the groundwork for the rational reengineering of NRPSs by swapping domains handling different substrates to generate novel natural products. Although many structural and biochemical studies of NRPSs exist, few studies have focused on the energetics and dynamics governing the interactions in these systems. Here, we present detailed binding studies of an adenylation domain and its partner carrier protein in apo-, holo-, and substrate-loaded forms. Results from fluorescence anisotropy, isothermal titration calorimetry, and NMR titrations indicated that covalent modifications to a carrier protein modulate domain communication, suggesting that chemical modifications to carrier proteins during NRPS synthesis may impart directionality to sequential NRPS domain interactions. Comparison of the structure and dynamics of an apo-aryl carrier protein with those of its modified forms revealed structural fluctuations induced by post-translational modifications and mediated by modulations of protein dynamics. The results provide a comprehensive molecular description of a carrier protein throughout its life cycle and demonstrate how a network of dynamic residues can propagate the molecular impact of chemical modifications throughout a protein and influence its affinity toward partner domains.
Keywords: adenylation domain; allostery; antibiotics; aryl carrier protein; nonribosomal peptide synthetase; nuclear magnetic resonance (NMR); protein dynamic; protein-protein interaction; structural biology.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures

Similar articles
-
Expression, purification, and characterization of HMWP2, a 229 kDa, six domain protein subunit of Yersiniabactin synthetase.Biochemistry. 2000 Apr 25;39(16):4729-39. doi: 10.1021/bi992923g. Biochemistry. 2000. PMID: 10769129
-
Purification, priming, and catalytic acylation of carrier protein domains in the polyketide synthase and nonribosomal peptidyl synthetase modules of the HMWP1 subunit of yersiniabactin synthetase.Proc Natl Acad Sci U S A. 2001 Jan 2;98(1):99-104. doi: 10.1073/pnas.98.1.99. Proc Natl Acad Sci U S A. 2001. PMID: 11134531 Free PMC article.
-
Analysis of the linker region joining the adenylation and carrier protein domains of the modular nonribosomal peptide synthetases.Proteins. 2014 Oct;82(10):2691-702. doi: 10.1002/prot.24635. Epub 2014 Jul 5. Proteins. 2014. PMID: 24975514 Free PMC article.
-
New Structural Data Reveal the Motion of Carrier Proteins in Nonribosomal Peptide Synthesis.Angew Chem Int Ed Engl. 2016 Aug 16;55(34):9834-40. doi: 10.1002/anie.201602614. Epub 2016 Jul 20. Angew Chem Int Ed Engl. 2016. PMID: 27435901 Free PMC article. Review.
-
[Advances in the study of the mechanism and application of nonribosomal peptide synthetases].Wei Sheng Wu Xue Bao. 2007 Aug;47(4):734-7. Wei Sheng Wu Xue Bao. 2007. PMID: 17944384 Review. Chinese.
Cited by
-
Using delayed decoupling to attenuate residual signals in editing filters.Magn Reson (Gott). 2021;2(1):475-487. doi: 10.5194/mr-2-475-2021. Epub 2021 Jun 21. Magn Reson (Gott). 2021. PMID: 34661195 Free PMC article.
-
Triepoxide formation by a flavin-dependent monooxygenase in monensin biosynthesis.Nat Commun. 2023 Oct 7;14(1):6273. doi: 10.1038/s41467-023-41889-0. Nat Commun. 2023. PMID: 37805629 Free PMC article.
-
NMR as a readout to monitor and restore the integrity of complex chemoenzymatic reactions.J Magn Reson. 2022 Sep;342:107265. doi: 10.1016/j.jmr.2022.107265. Epub 2022 Jul 8. J Magn Reson. 2022. PMID: 35849973 Free PMC article.
-
Probing Substrate-Loaded Carrier Proteins by Nuclear Magnetic Resonance.Methods Mol Biol. 2023;2670:235-253. doi: 10.1007/978-1-0716-3214-7_12. Methods Mol Biol. 2023. PMID: 37184708
-
Finding Druggable Sites in Proteins Using TACTICS.J Chem Inf Model. 2021 Jun 28;61(6):2897-2910. doi: 10.1021/acs.jcim.1c00204. Epub 2021 Jun 7. J Chem Inf Model. 2021. PMID: 34096704 Free PMC article.
References
-
- Walsh C. T. (2003) Antibiotics: Actions, Origins, Resistance, 1st Ed., ASM Press, Washington, D. C.
-
- Fischbach M. A., and Walsh C. T. (2006) Assembly-line enzymology for polyketide and nonribosomal peptide antibiotics: logic, machinery, and mechanisms. Chem. Rev. 106, 3468–3496 - PubMed
-
- Marahiel M. A., Stachelhaus T., and Mootz H. D. (1997) Modular peptide synthetases involved in nonribosomal peptide synthesis. Chem. Rev. 97, 2651–2674 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
