

Catalytic oxidant scavenging by selenium-containing compounds: Reduction of selenoxides and N-chloramines by thiols and redox enzymes
- PMID: 28458184
- PMCID: PMC5408155
- DOI: 10.1016/j.redox.2017.04.023
Catalytic oxidant scavenging by selenium-containing compounds: Reduction of selenoxides and N-chloramines by thiols and redox enzymes
Abstract
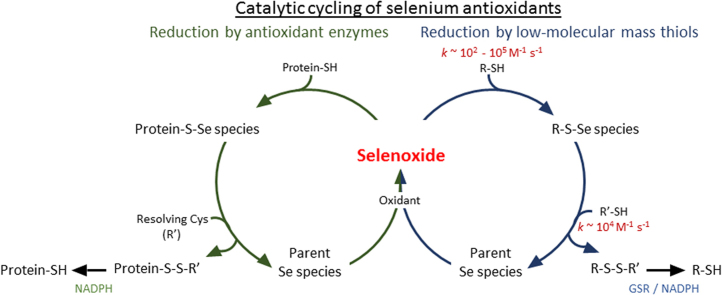
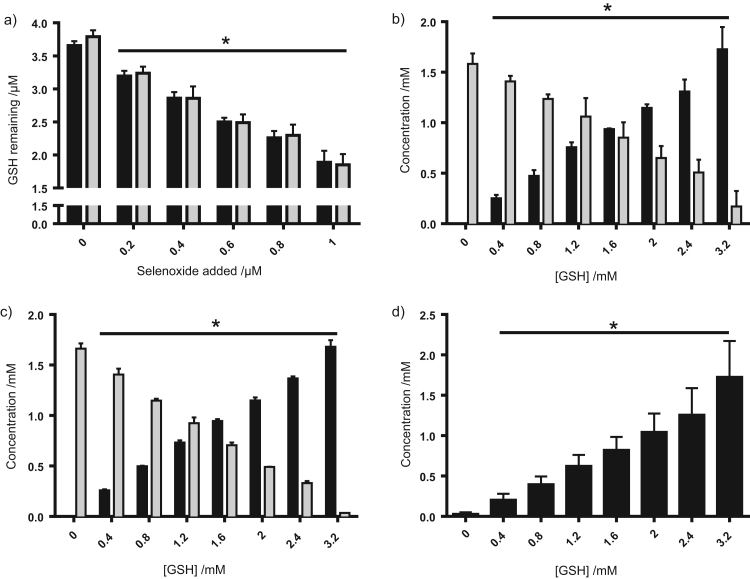
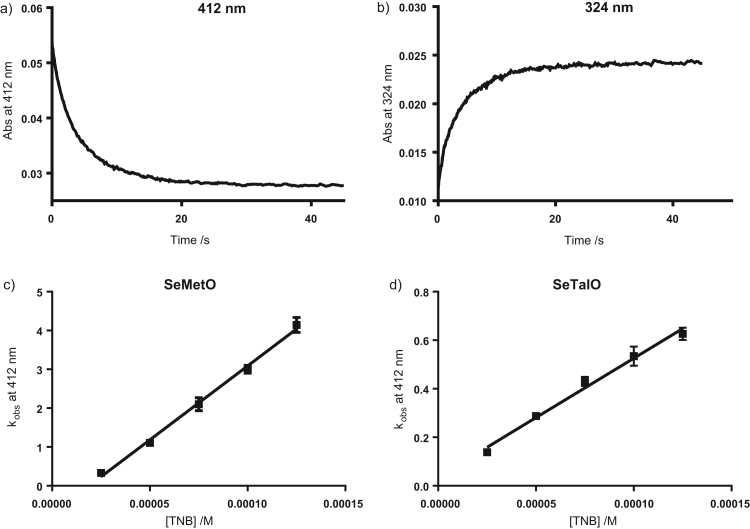
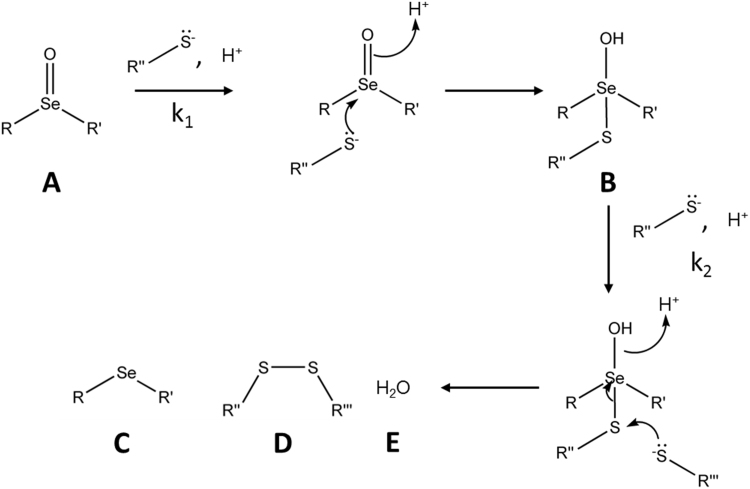
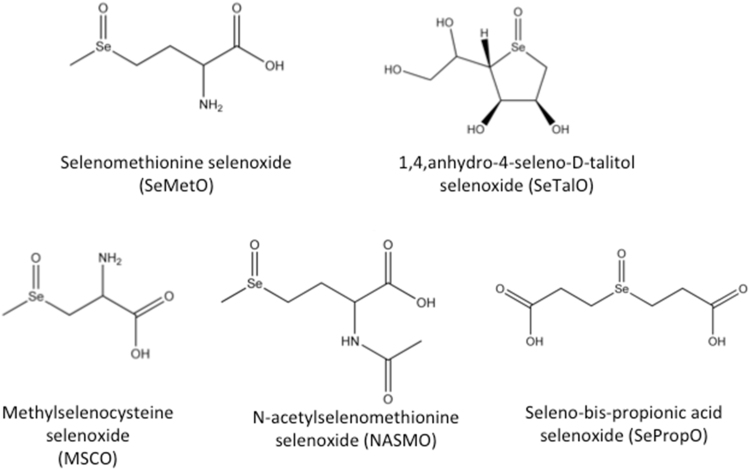
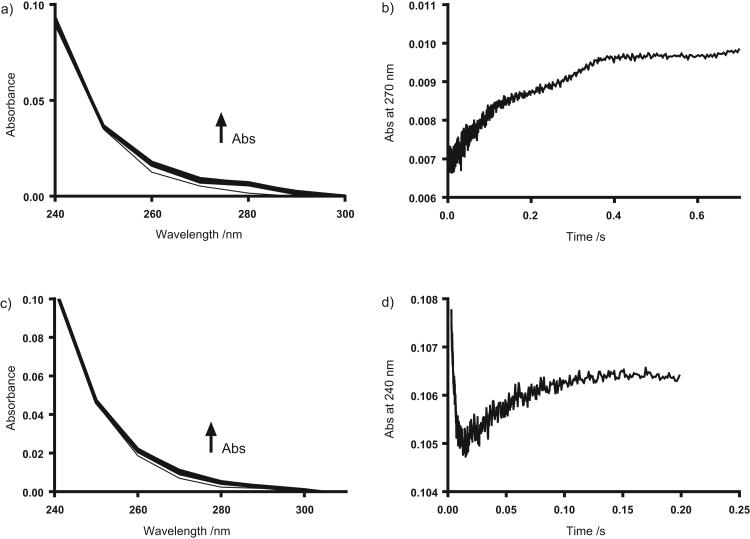
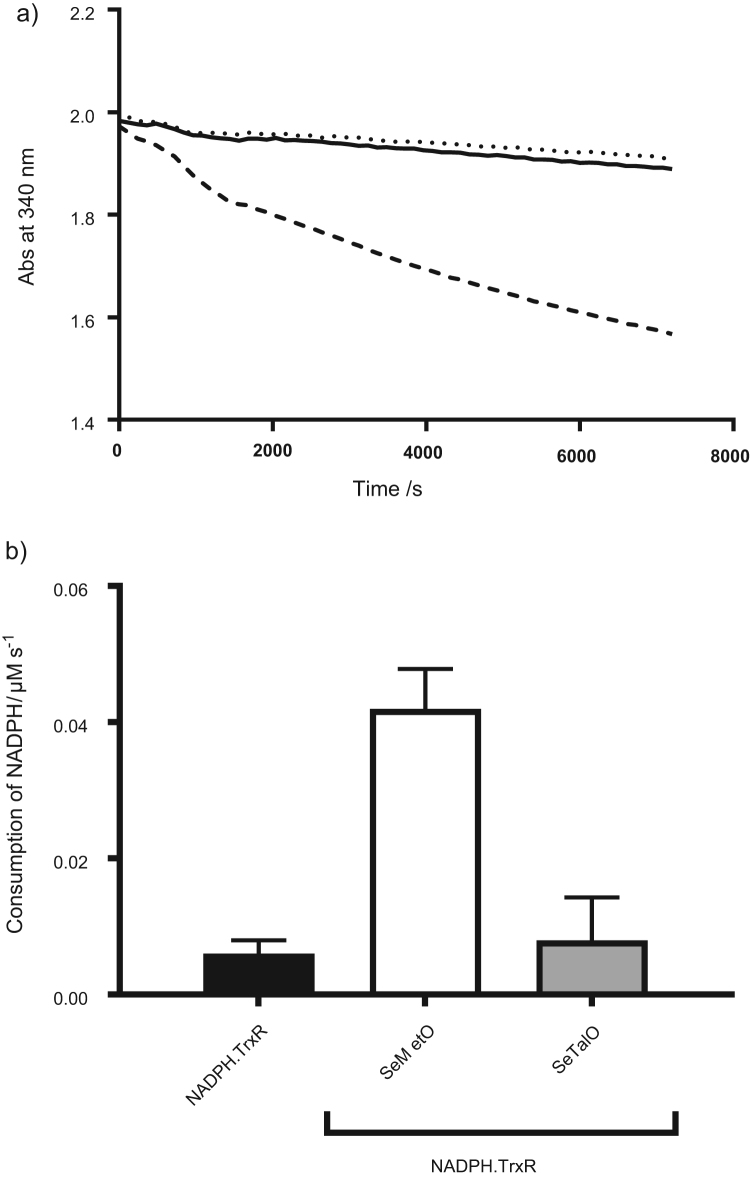
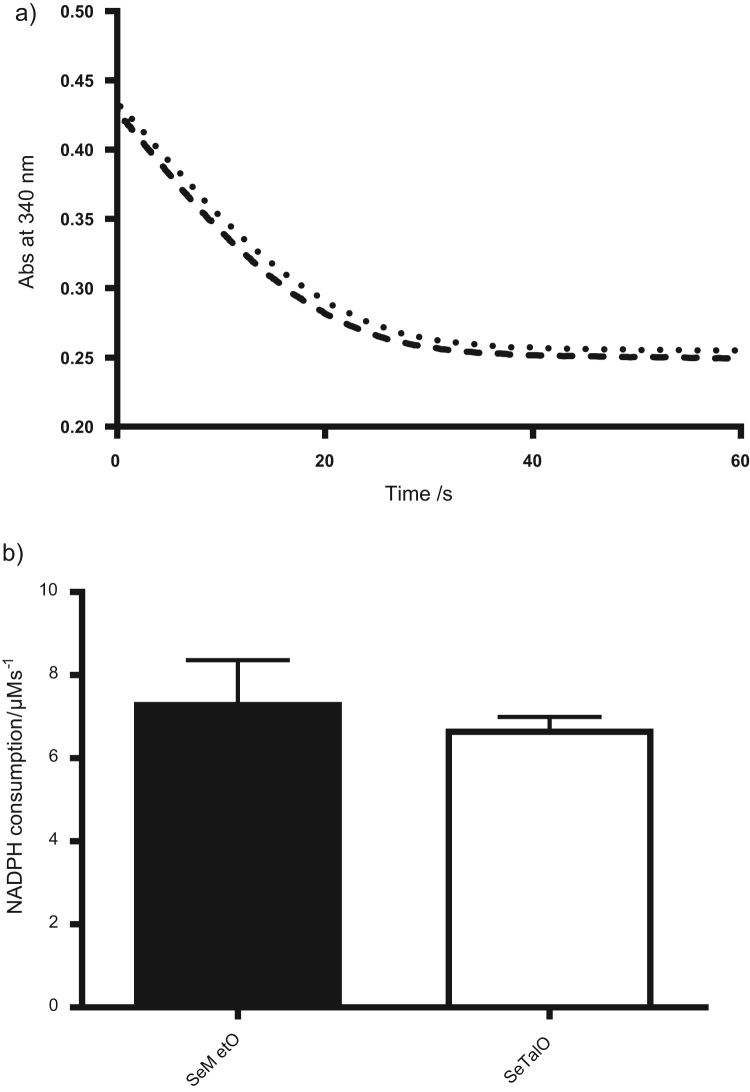
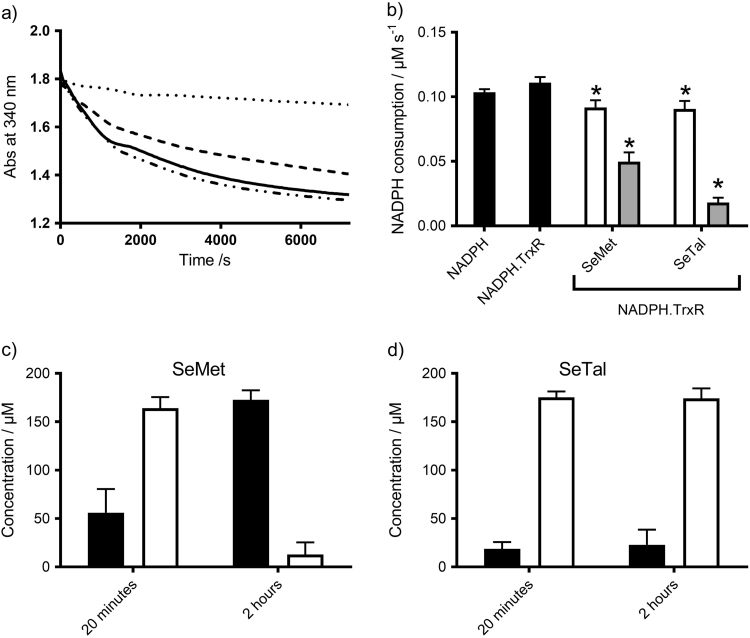
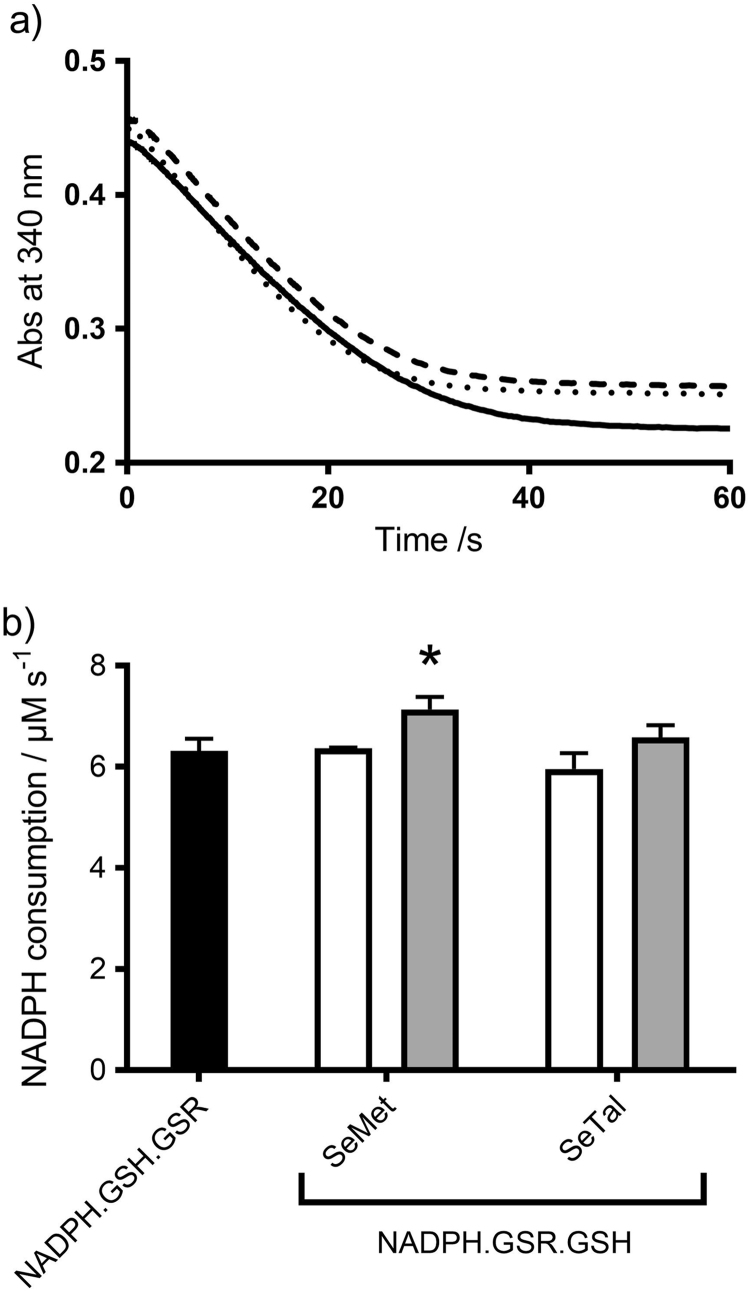
Myeloperoxidase produces strong oxidants during the immune response to destroy invading pathogens. However, these oxidants can also cause tissue damage, which contributes to the development of numerous inflammatory diseases. Selenium containing compounds, including selenomethionine (SeMet) and 1,4-anhydro-5-seleno-D-talitol (SeTal), react rapidly with different MPO-derived oxidants to form the respective selenoxides (SeMetO and SeTalO). This study investigates the susceptibility of these selenoxides to undergo reduction back to the parent compounds by intracellular reducing systems, including glutathione (GSH) and the glutathione reductase and thioredoxin reductase systems. GSH is shown to reduce SeMetO and SeTalO, with consequent formation of GSSG with apparent second order rate constants, k2, in the range 103-104M-1s-1. Glutathione reductase reduces both SeMetO and SeTalO at the expense of NADPH via formation of GSSG, whereas thioredoxin reductase acts only on SeMetO. The presence of SeMet and SeTal also increased the rate at which NADPH was consumed by the glutathione reductase system in the presence of N-chloramines. In contrast, the presence of SeMet and SeTal reduced the rate of NADPH consumption by the thioredoxin reductase system after addition of N-chloramines, consistent with the rapid formation of selenoxides, but only slow reduction by thioredoxin reductase. These results support a potential role of seleno compounds to act as catalytic scavengers of MPO-derived oxidants, particularly in the presence of glutathione reductase and NADPH, assuming that sufficient plasma levels of the parent selenoether can be achieved in vivo following supplementation.
Keywords: Antioxidants; Glutathione; Glutathione reductase; Hypochlorous acid; Myeloperoxidase; N-chloramines; Selenium; Thioredoxin reductase.
Copyright © 2017. Published by Elsevier B.V.
Figures
References
-
- Davies M.J., Hawkins C.L., Pattison D.I., Rees M.D. Mammalian heme peroxidases: from molecular mechanisms to health implications. Antioxid. Redox Signal. 2008;10:1199–1234. - PubMed
-
- Hampton M.B., Kettle A.J., Winterbourn C.C. Inside the neutrophil phagosome: oxidants, myeloperoxidase, and bacterial killing. Blood. 1998;92:3007–3017. - PubMed
-
- Rayner B.S., Love D.T., Hawkins C.L. Comparative reactivity of myeloperoxidase-derived oxidants with mammalian cells. Free Radic. Biol. Med. 2014;71:240–255. - PubMed
-
- Thomas E.L., Grisham M.B., Jefferson M.M. Preparation and characterization of chloramines. Methods Enzymol. 1986;132:569–585. - PubMed
-
- Klebanoff S.J. Myeloperoxidase: friend and foe. J. Leukoc. Biol. 2005;77:598–625. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
