

Conservation and innovation in the DUX4-family gene network
- PMID: 28459454
- PMCID: PMC5446306
- DOI: 10.1038/ng.3846
Conservation and innovation in the DUX4-family gene network
Abstract
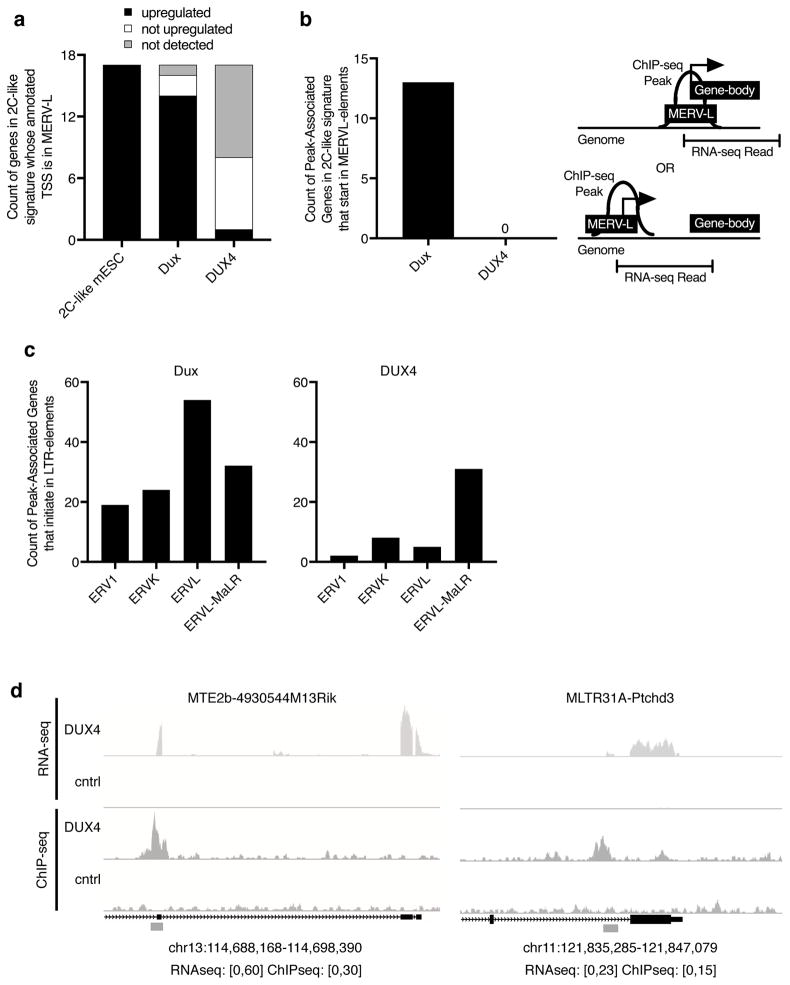
Facioscapulohumeral dystrophy (FSHD; MIM158900, MIM158901) is caused by misexpression of the DUX4 transcription factor in skeletal muscle. Animal models of FSHD are hindered by incomplete knowledge regarding the conservation of the DUX4 transcriptional program in other species. Despite the divergence of their binding motifs, both mouse DUX and human DUX4 in mouse and human muscle cells, respectively, activate genes associated with cleavage-stage embryos, including MERVL and ERVL-MaLR retrotransposons. We found that human DUX4 expressed in mouse cells maintained modest activation of cleavage-stage genes driven by conventional promoters but did not activate MERVL-promoted genes. Thus, the ancestral DUX4-regulated genes are characteristic of cleavage-stage embryos and are driven by conventional promoters, whereas divergence of the DUX4 and DUX homeodomains correlates with retrotransposon specificity. These results provide insight into how species balance conservation of a core transcriptional program with innovation at retrotransposon promoters, and establish a basis for animal models recreating the FSHD transcriptome.
Figures




References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
