

DNA sequence homology induces cytosine-to-thymine mutation by a heterochromatin-related pathway in Neurospora
- PMID: 28459455
- PMCID: PMC5474309
- DOI: 10.1038/ng.3857
DNA sequence homology induces cytosine-to-thymine mutation by a heterochromatin-related pathway in Neurospora
Abstract
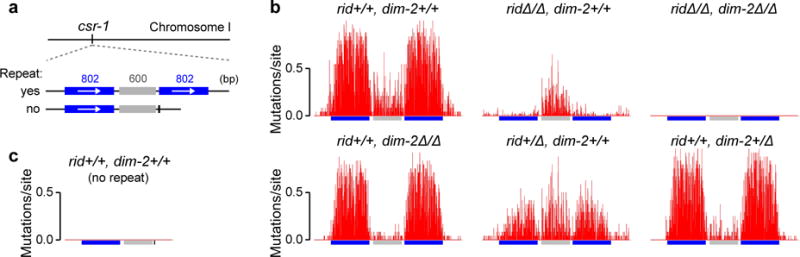
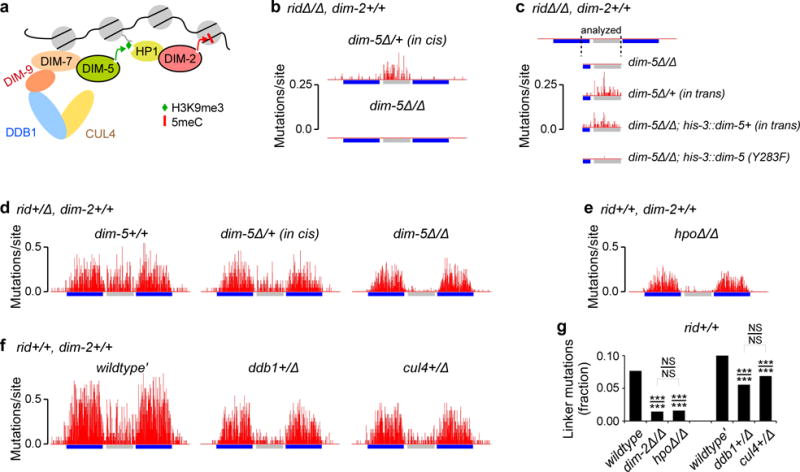
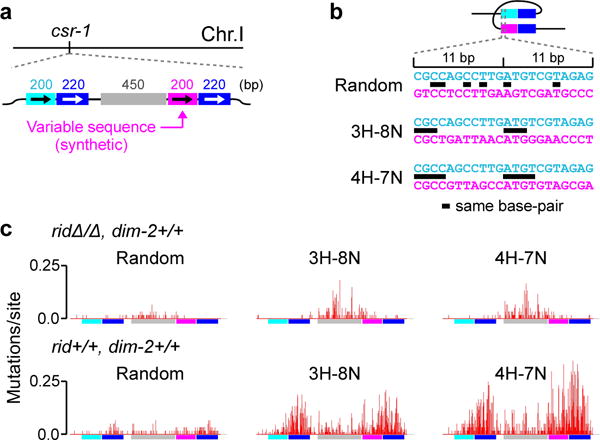
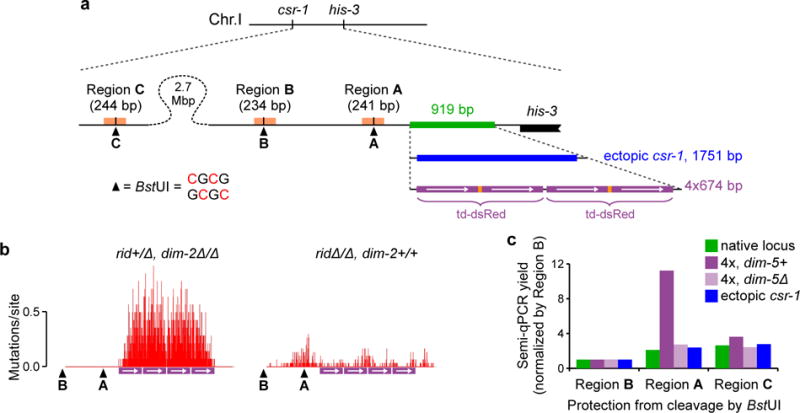
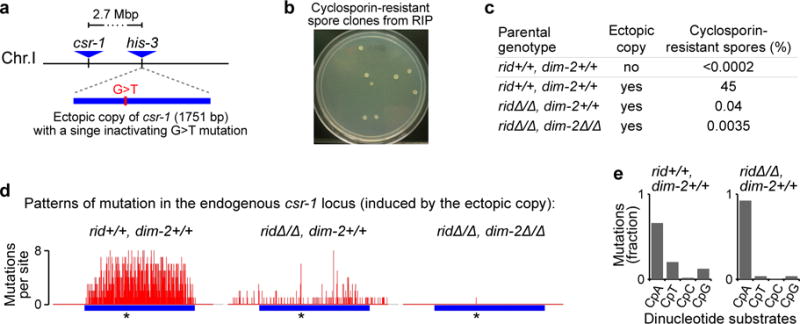
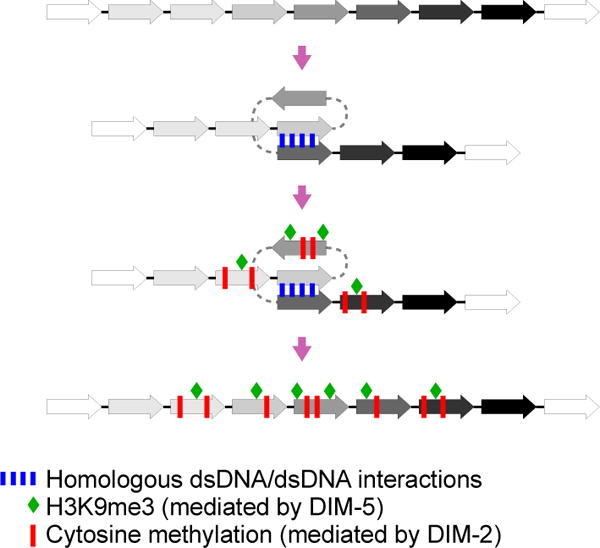
Most eukaryotic genomes contain substantial amounts of repetitive DNA organized in the form of constitutive heterochromatin and associated with repressive epigenetic modifications, such as H3K9me3 and C5 cytosine methylation (5mC). In the fungus Neurospora crassa, H3K9me3 and 5mC are catalyzed, respectively, by a conserved SUV39 histone methyltransferase, DIM-5, and a DNMT1-like cytosine methyltransferase, DIM-2. Here we show that DIM-2 can also mediate repeat-induced point mutation (RIP) of repetitive DNA in N. crassa. We further show that DIM-2-dependent RIP requires DIM-5, HP1, and other known heterochromatin factors, implying a role for a repeat-induced heterochromatin-related process. Our previous findings suggest that the mechanism of repeat recognition for RIP involves direct interactions between homologous double-stranded DNA (dsDNA) segments. We thus now propose that, in somatic cells, homologous dsDNA-dsDNA interactions between a small number of repeat copies can nucleate a transient heterochromatic state, which, on longer repeat arrays, may lead to the formation of constitutive heterochromatin.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
Relics of repeat-induced point mutation direct heterochromatin formation in Neurospora crassa.Genome Res. 2009 Mar;19(3):427-37. doi: 10.1101/gr.086231.108. Epub 2008 Dec 17. Genome Res. 2009. PMID: 19092133 Free PMC article.
-
Induction of H3K9me3 and DNA methylation by tethered heterochromatin factors in Neurospora crassa.Proc Natl Acad Sci U S A. 2017 Nov 7;114(45):E9598-E9607. doi: 10.1073/pnas.1715049114. Epub 2017 Oct 23. Proc Natl Acad Sci U S A. 2017. PMID: 29078403 Free PMC article.
-
DNA methylation and the formation of heterochromatin in Neurospora crassa.Heredity (Edinb). 2010 Jul;105(1):38-44. doi: 10.1038/hdy.2010.44. Epub 2010 Apr 21. Heredity (Edinb). 2010. PMID: 20407471 Review.
-
Neurospora importin α is required for normal heterochromatic formation and DNA methylation.PLoS Genet. 2015 Mar 20;11(3):e1005083. doi: 10.1371/journal.pgen.1005083. eCollection 2015 Mar. PLoS Genet. 2015. PMID: 25793375 Free PMC article.
-
Recombination-independent recognition of DNA homology for repeat-induced point mutation.Curr Genet. 2017 Jun;63(3):389-400. doi: 10.1007/s00294-016-0649-4. Epub 2016 Sep 14. Curr Genet. 2017. PMID: 27628707 Free PMC article. Review.
Cited by
-
Rad54 Drives ATP Hydrolysis-Dependent DNA Sequence Alignment during Homologous Recombination.Cell. 2020 Jun 11;181(6):1380-1394.e18. doi: 10.1016/j.cell.2020.04.056. Epub 2020 Jun 4. Cell. 2020. PMID: 32502392 Free PMC article.
-
Remodeling of perturbed chromatin can initiate de novo transcriptional and post-transcriptional silencing.Proc Natl Acad Sci U S A. 2024 Jul 30;121(31):e2402944121. doi: 10.1073/pnas.2402944121. Epub 2024 Jul 25. Proc Natl Acad Sci U S A. 2024. PMID: 39052837 Free PMC article.
-
A combination of transcription factors mediates inducible interchromosomal contacts.Elife. 2019 May 13;8:e42499. doi: 10.7554/eLife.42499. Elife. 2019. PMID: 31081754 Free PMC article.
-
Intragenomic rDNA variation - the product of concerted evolution, mutation, or something in between?Heredity (Edinb). 2023 Sep;131(3):179-188. doi: 10.1038/s41437-023-00634-5. Epub 2023 Jul 4. Heredity (Edinb). 2023. PMID: 37402824 Free PMC article. Review.
-
RID is required for both repeat-induced point mutation and nucleation of a novel transitional heterochromatic state for euchromatic repeats.Nucleic Acids Res. 2025 Mar 20;53(6):gkaf263. doi: 10.1093/nar/gkaf263. Nucleic Acids Res. 2025. PMID: 40183634 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
