

Conserved roles of mouse DUX and human DUX4 in activating cleavage-stage genes and MERVL/HERVL retrotransposons
- PMID: 28459457
- PMCID: PMC5703070
- DOI: 10.1038/ng.3844
Conserved roles of mouse DUX and human DUX4 in activating cleavage-stage genes and MERVL/HERVL retrotransposons
Abstract
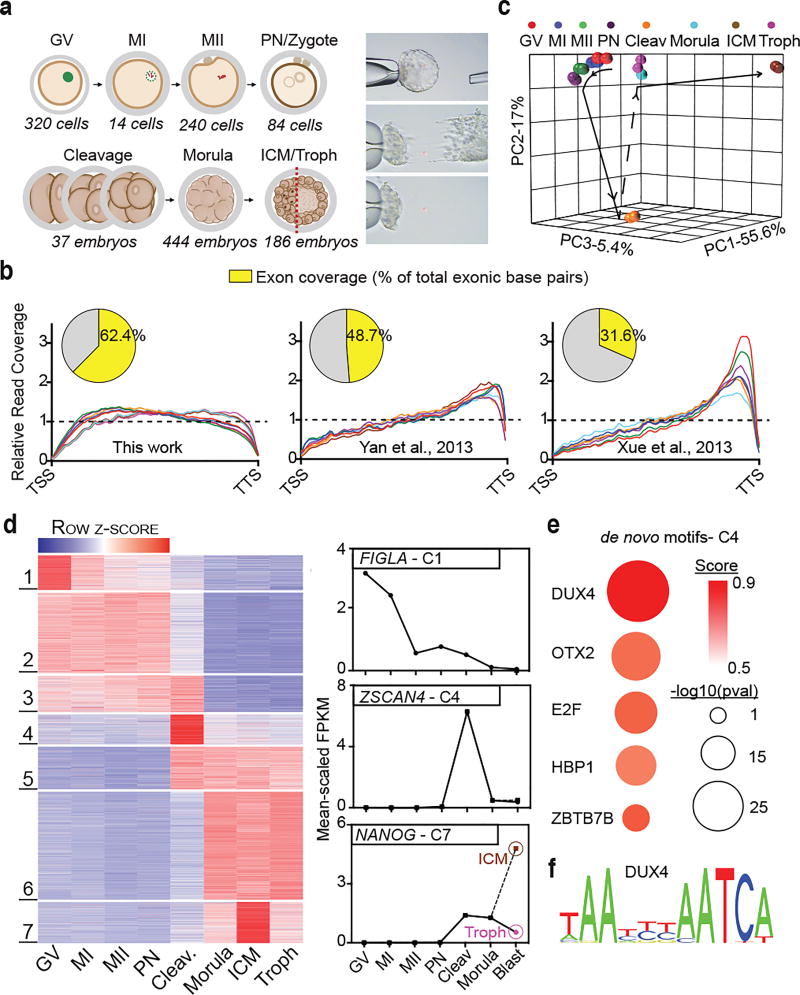
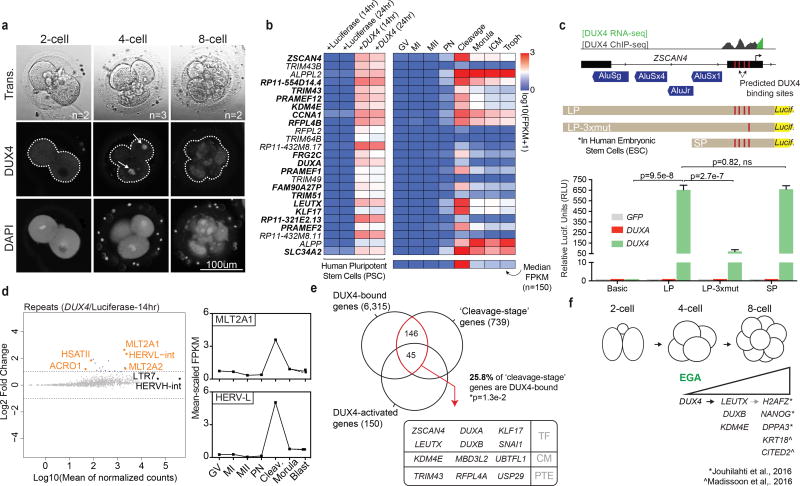
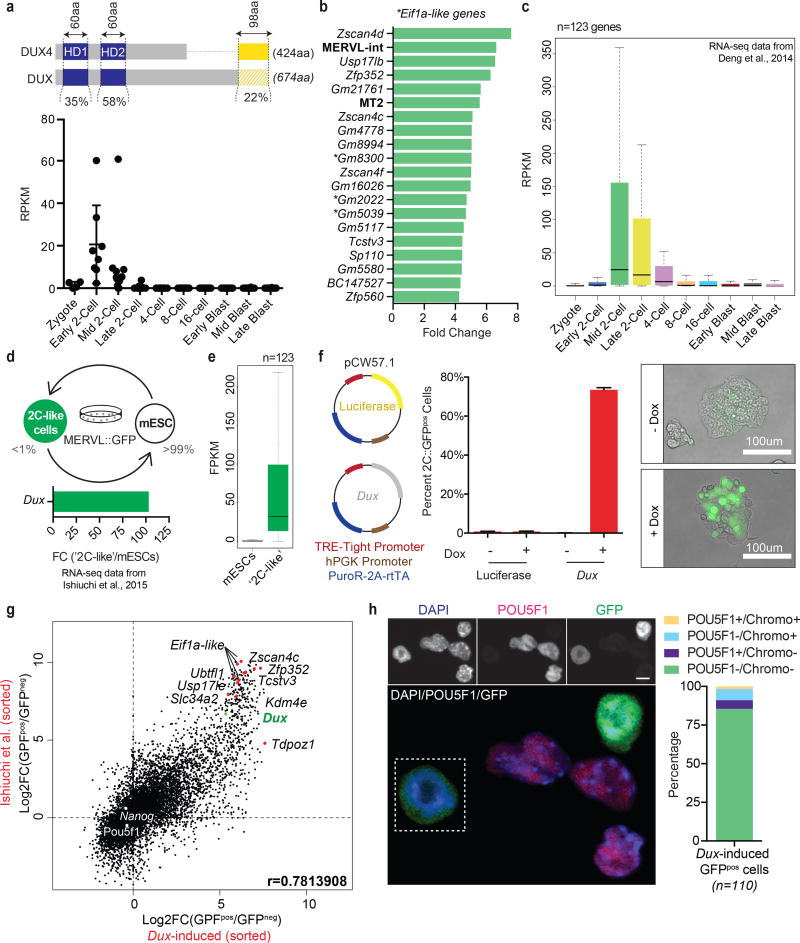
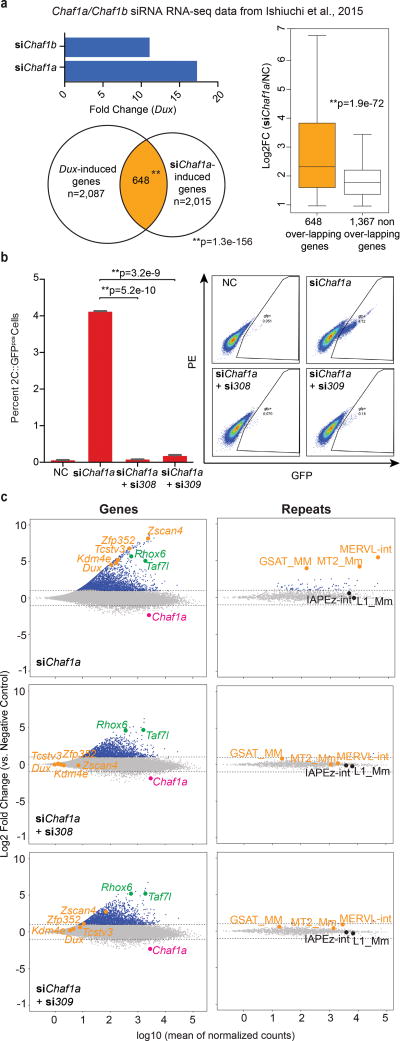
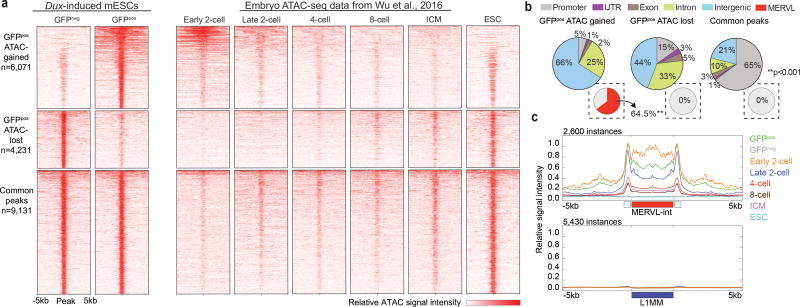
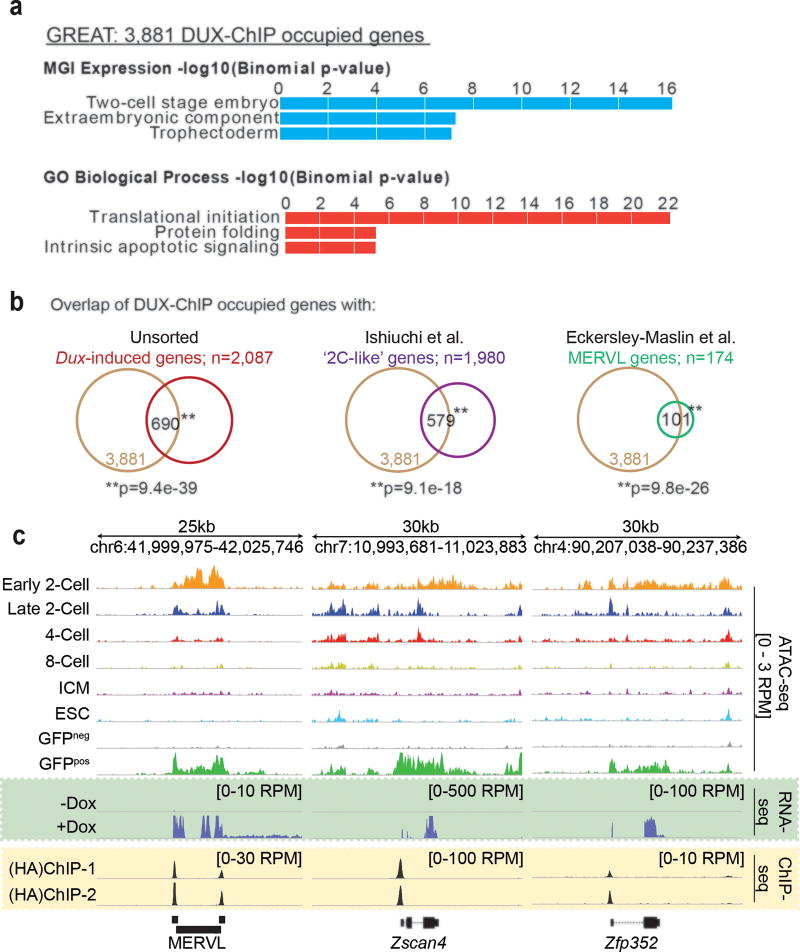
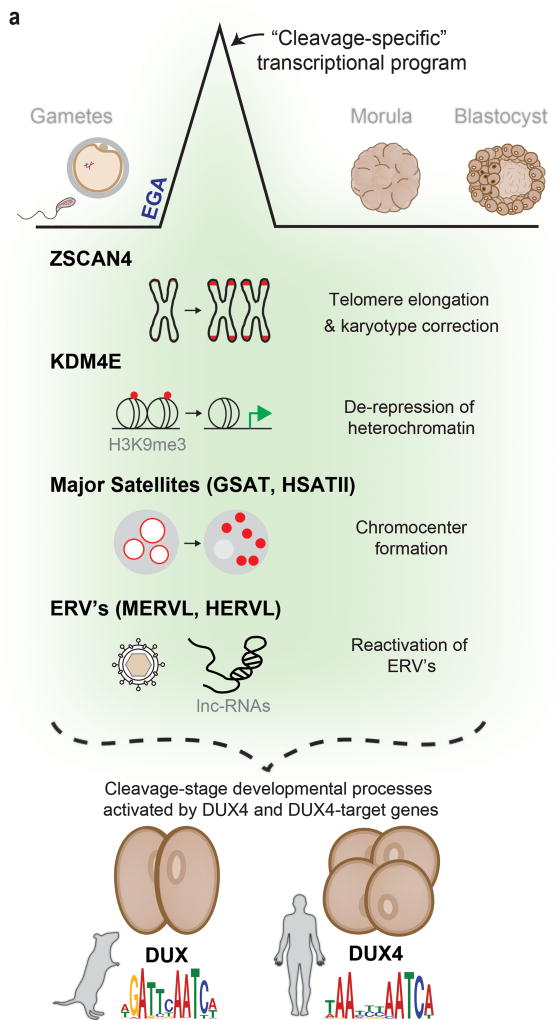
To better understand transcriptional regulation during human oogenesis and preimplantation development, we defined stage-specific transcription, which highlighted the cleavage stage as being highly distinctive. Here, we present multiple lines of evidence that a eutherian-specific multicopy retrogene, DUX4, encodes a transcription factor that activates hundreds of endogenous genes (for example, ZSCAN4, KDM4E and PRAMEF-family genes) and retroviral elements (MERVL/HERVL family) that define the cleavage-specific transcriptional programs in humans and mice. Remarkably, mouse Dux expression is both necessary and sufficient to convert mouse embryonic stem cells (mESCs) into 2-cell-embryo-like ('2C-like') cells, measured here by the reactivation of '2C' genes and repeat elements, the loss of POU5F1 (also known as OCT4) protein and chromocenters, and the conversion of the chromatin landscape (as assessed by transposase-accessible chromatin using sequencing (ATAC-seq)) to a state strongly resembling that of mouse 2C embryos. Thus, we propose mouse DUX and human DUX4 as major drivers of the cleavage or 2C state.
Conflict of interest statement
The authors have no competing financial interests.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
