

Molecular and Functional Bases of Selection against a Mutation Bias in an RNA Virus
- PMID: 28460010
- PMCID: PMC5433387
- DOI: 10.1093/gbe/evx075
Molecular and Functional Bases of Selection against a Mutation Bias in an RNA Virus
Abstract
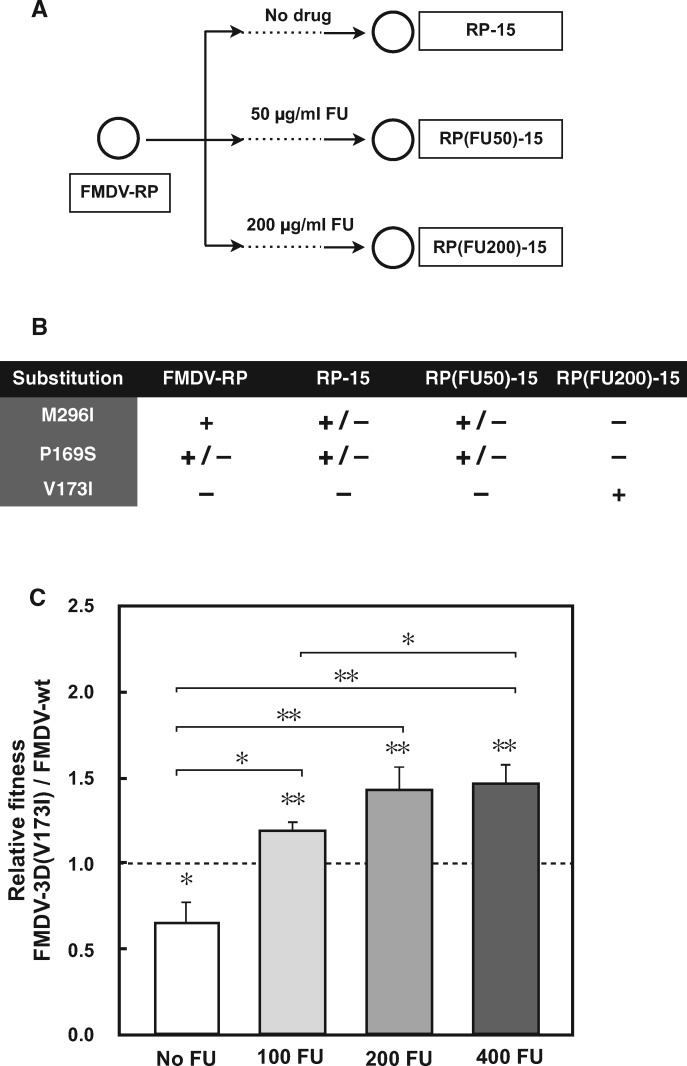
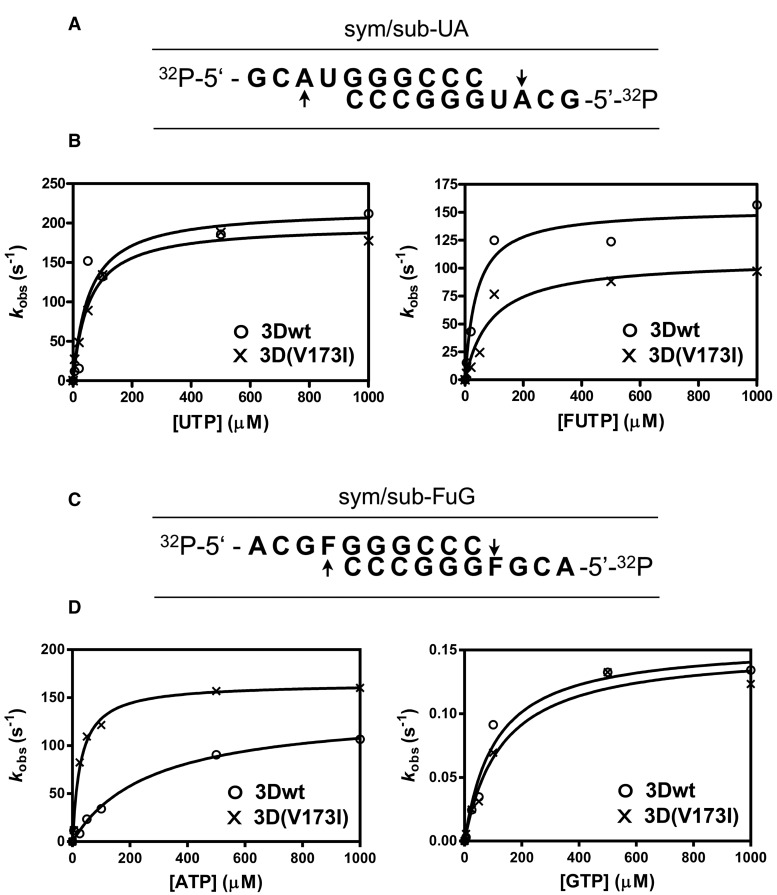
The selective pressures acting on viruses that replicate under enhanced mutation rates are largely unknown. Here, we describe resistance of foot-and-mouth disease virus to the mutagen 5-fluorouracil (FU) through a single polymerase substitution that prevents an excess of A to G and U to C transitions evoked by FU on the wild-type foot-and-mouth disease virus, while maintaining the same level of mutant spectrum complexity. The polymerase substitution inflicts upon the virus a fitness loss during replication in absence of FU but confers a fitness gain in presence of FU. The compensation of mutational bias was documented by in vitro nucleotide incorporation assays, and it was associated with structural modifications at the N-terminal region and motif B of the viral polymerase. Predictions of the effect of mutations that increase the frequency of G and C in the viral genome and encoded polymerase suggest multiple points in the virus life cycle where the mutational bias in favor of G and C may be detrimental. Application of predictive algorithms suggests adverse effects of the FU-directed mutational bias on protein stability. The results reinforce modulation of nucleotide incorporation as a lethal mutagenesis-escape mechanism (that permits eluding virus extinction despite replication in the presence of a mutagenic agent) and suggest that mutational bias can be a target of selection during virus replication.
Keywords: 5-fluorouracil; antiviral resistance; fitness; foot-and-mouth disease virus; lethal mutagenesis; protein folding stability.
© The Author 2017. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures
References
-
- Agudo R, Arias A, Domingo E.. 2009. 5-Fluorouracil in lethal mutagenesis of foot-and-mouth disease virus. Future Med Chem. 1:529–539. - PubMed
-
- Agudo R, et al.2008. Molecular characterization of a dual inhibitory and mutagenic activity of 5-fluorouridine triphosphate on viral RNA synthesis. Implications for lethal mutagenesis. J Mol Biol. 382:652–666. - PubMed
-
- Airaksinen A, Pariente N, Menendez-Arias L, Domingo E.. 2003. Curing of foot-and-mouth disease virus from persistently infected cells by ribavirin involves enhanced mutagenesis. Virology 311:339–349. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
