

Triptolide suppresses the in vitro and in vivo growth of lung cancer cells by targeting hyaluronan-CD44/RHAMM signaling
- PMID: 28460475
- PMCID: PMC5432308
- DOI: 10.18632/oncotarget.15879
Triptolide suppresses the in vitro and in vivo growth of lung cancer cells by targeting hyaluronan-CD44/RHAMM signaling
Abstract
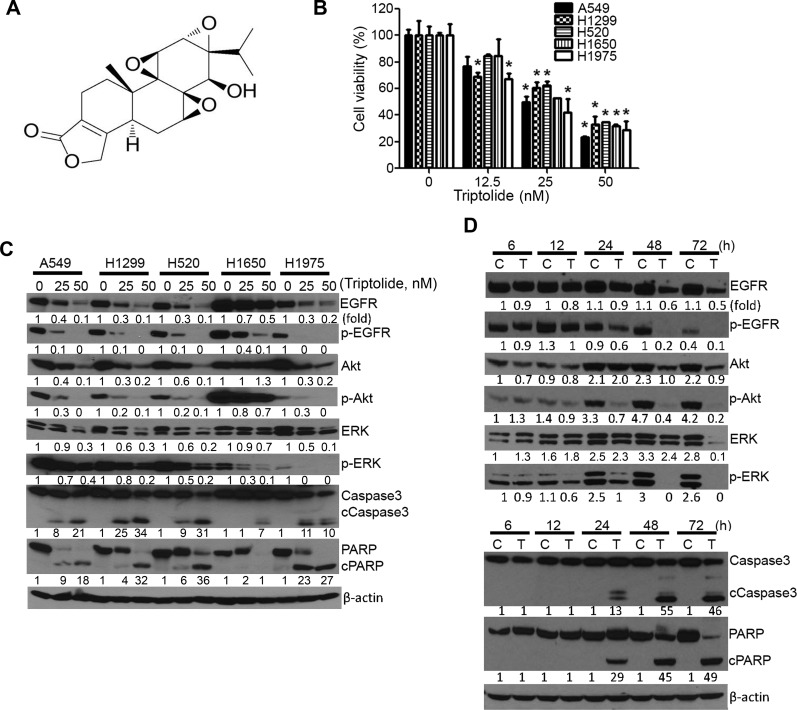
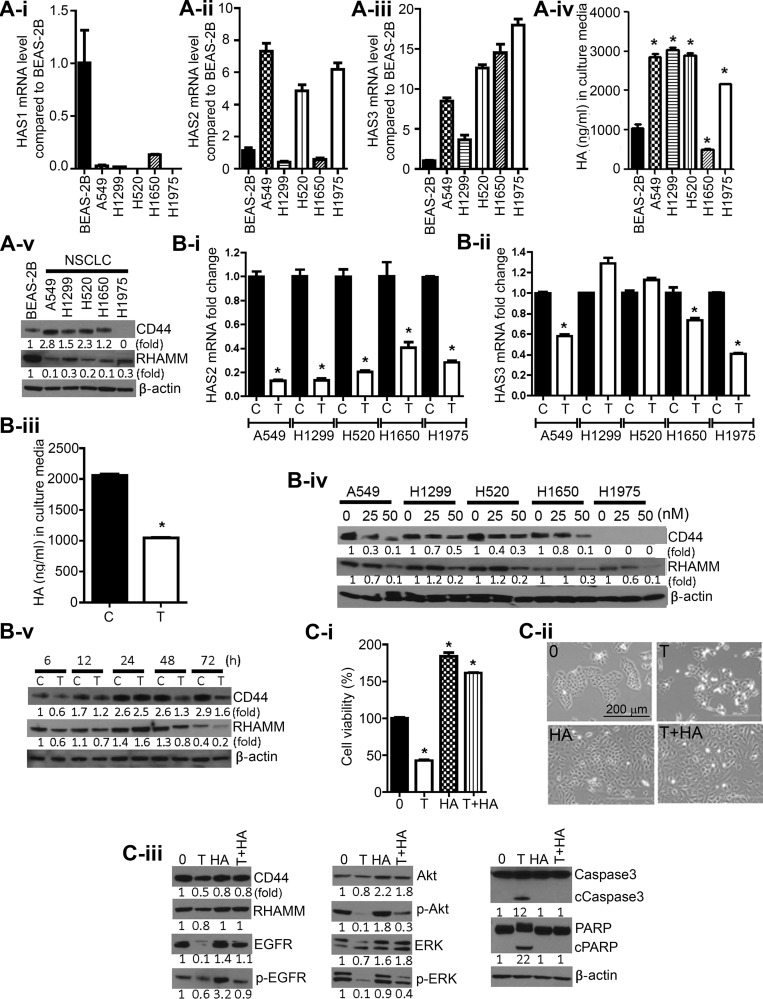
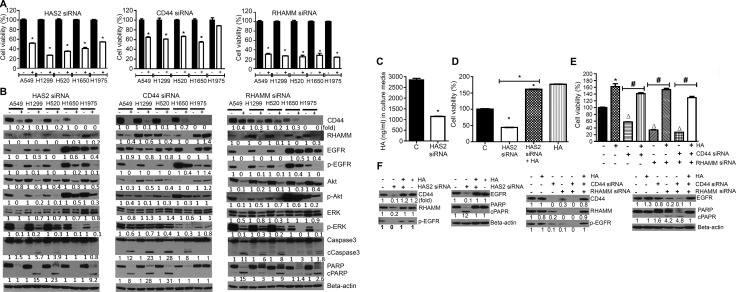
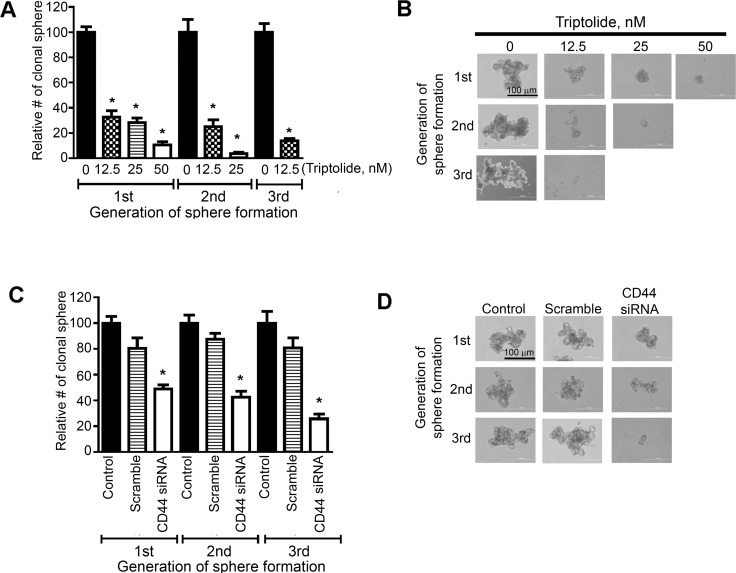
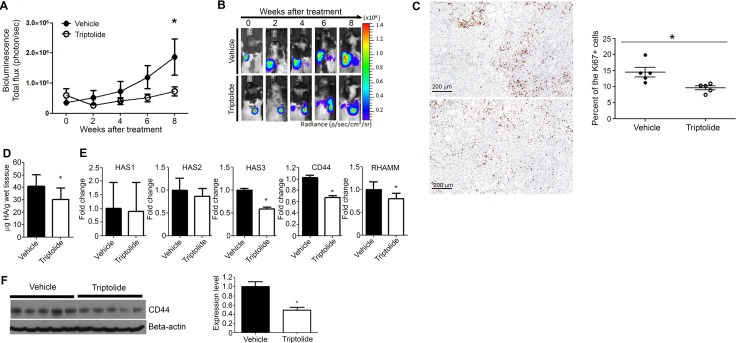
Higher levels of hyaluronan (HA) and its receptors CD44 and RHAMM have been associated with poor prognosis and metastasis in NSCLC. In the current study, our goal was to define, using cellular and orthotopic lung tumor models, the role of HA-CD44/RHAMM signaling in lung carcinogenesis and to assess the potential of triptolide to block HA-CD44/RHAMM signaling and thereby suppress the development and progression of lung cancer. Triptolide reduced the viability of five non-small cell lung cancer (NSCLC) cells, the proliferation and self-renewal of pulmospheres, and levels of HA synthase 2 (HAS2), HAS3, HA, CD44, RHAMM, EGFR, Akt and ERK, but increased the cleavage of caspase 3 and PARP. Silencing of HAS2, CD44 or RHAMM induced similar effects. Addition of excess HA to the culture media completely abrogated the effects of triptolide and siRNAs targeting HAS2, CD44, or RHAMM. In an orthotopic lung cancer model in nude rats, intranasal administration of liposomal triptolide (400 μg/kg) for 8 weeks significantly reduced lung tumor growth as determined by bioluminescence imaging, lung weight measurements and gross and histopathological analysis of tumor burden. Also, triptolide suppressed expressions of Ki-67, a marker for cell proliferation, HAS2, HAS3, HA, CD44, and RHAMM in lung tumors. Overall, our results provide a strong rationale for mitigating lung cancer by targeting the HA-CD44/RHAMM signaling axis.
Keywords: NSCLC; hyaluronan; hyaluronan receptor; orthotopic; triptolide.
Conflict of interest statement
We confirm that there are no conflicts of interest associated with this publication.
Figures
Similar articles
-
Hyaluronan-CD44/RHAMM interaction-dependent cell proliferation and survival in lung cancer cells.Mol Carcinog. 2019 Mar;58(3):321-333. doi: 10.1002/mc.22930. Epub 2018 Nov 12. Mol Carcinog. 2019. PMID: 30365189 Free PMC article.
-
Oligosaccharides of hyaluronan induce angiogenesis through distinct CD44 and RHAMM-mediated signalling pathways involving Cdc2 and gamma-adducin.Int J Oncol. 2009 Oct;35(4):761-73. doi: 10.3892/ijo_00000389. Int J Oncol. 2009. PMID: 19724912
-
The impact of the receptor of hyaluronan-mediated motility (RHAMM) on human urothelial transitional cell cancer of the bladder.PLoS One. 2013 Sep 17;8(9):e75681. doi: 10.1371/journal.pone.0075681. eCollection 2013. PLoS One. 2013. PMID: 24069434 Free PMC article.
-
Roles and targeting of the HAS/hyaluronan/CD44 molecular system in cancer.Matrix Biol. 2017 May;59:3-22. doi: 10.1016/j.matbio.2016.10.001. Epub 2016 Oct 13. Matrix Biol. 2017. PMID: 27746219 Review.
-
Interactions between Hyaluronan and Its Receptors (CD44, RHAMM) Regulate the Activities of Inflammation and Cancer.Front Immunol. 2015 May 6;6:201. doi: 10.3389/fimmu.2015.00201. eCollection 2015. Front Immunol. 2015. PMID: 25999946 Free PMC article. Review.
Cited by
-
An Overview of Saturated Cyclic Ethers: Biological Profiles and Synthetic Strategies.Molecules. 2019 Oct 21;24(20):3778. doi: 10.3390/molecules24203778. Molecules. 2019. PMID: 31640154 Free PMC article. Review.
-
Triptolide inhibits angiogenesis in microvascular endothelial cells through regulation of miR-92a.J Physiol Biochem. 2019 Nov;75(4):573-583. doi: 10.1007/s13105-019-00707-2. Epub 2019 Nov 5. J Physiol Biochem. 2019. Retraction in: J Physiol Biochem. 2022 Aug;78(3):705. doi: 10.1007/s13105-022-00879-4. PMID: 31691162 Retracted.
-
IGFBP-3 Blocks Hyaluronan-CD44 Signaling, Leading to Increased Acetylcholinesterase Levels in A549 Cell Media and Apoptosis in a p53-Dependent Manner.Sci Rep. 2020 Mar 19;10(1):5083. doi: 10.1038/s41598-020-61743-3. Sci Rep. 2020. PMID: 32193421 Free PMC article.
-
A Mechanistic Overview of Triptolide and Celastrol, Natural Products from Tripterygium wilfordii Hook F.Front Pharmacol. 2018 Feb 14;9:104. doi: 10.3389/fphar.2018.00104. eCollection 2018. Front Pharmacol. 2018. PMID: 29491837 Free PMC article. Review.
-
Inhibition of lung adenocarcinoma by combinations of sulfasalazine (SAS) and disulfiram-copper (DSF-Cu) in cell line models and mice.Carcinogenesis. 2023 Jun 24;44(4):291-303. doi: 10.1093/carcin/bgad020. Carcinogenesis. 2023. PMID: 37053033 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
