

Characterization of the Zika virus two-component NS2B-NS3 protease and structure-assisted identification of allosteric small-molecule antagonists
- PMID: 28461069
- PMCID: PMC5558795
- DOI: 10.1016/j.antiviral.2017.04.015
Characterization of the Zika virus two-component NS2B-NS3 protease and structure-assisted identification of allosteric small-molecule antagonists
Abstract
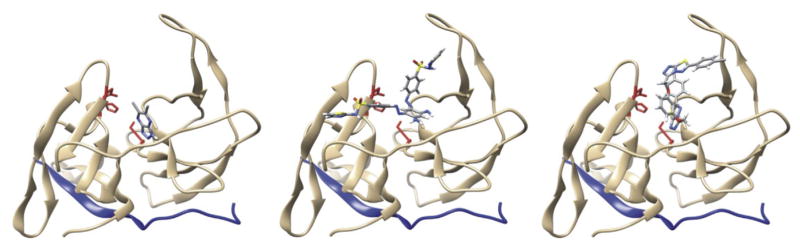
The recent re-emergence of Zika virus (ZIKV)1, a member of the Flaviviridae family, has become a global emergency. Currently, there are no effective methods of preventing or treating ZIKV infection, which causes severe neuroimmunopathology and is particularly harmful to the developing fetuses of infected pregnant women. However, the pathology induced by ZIKV is unique among flaviviruses, and knowledge of the biology of other family members cannot easily be extrapolated to ZIKV. Thus, structure-function studies of ZIKV proteins are urgently needed to facilitate the development of effective preventative and therapeutic agents. Like other flaviviruses, ZIKV expresses an NS2B-NS3 protease, which consists of the NS2B cofactor and the NS3 protease domain and is essential for cleavage of the ZIKV polyprotein precursor and generation of fully functional viral proteins. Here, we report the enzymatic characterization of ZIKV protease, and we identify structural scaffolds for allosteric small-molecule inhibitors of this protease. Molecular modeling of the protease-inhibitor complexes suggests that these compounds bind to the druggable cavity in the NS2B-NS3 protease interface and affect productive interactions of the protease domain with its cofactor. The most potent compound demonstrated efficient inhibition of ZIKV propagation in vitro in human fetal neural progenitor cells and in vivo in SJL mice. The inhibitory scaffolds could be further developed into valuable research reagents and, ultimately, provide a roadmap for the selection of efficient inhibitors of ZIKV infection.
Keywords: Flavivirus; Inhibitors; NS2B; NS3; Protease; Zika virus.
Copyright © 2017 Elsevier B.V. All rights reserved.
Conflict of interest statement
No author has an actual or perceived conflict of interest with the contents of this article.
Figures






References
-
- Azevedo RS, Araujo MT, Martins Filho AJ, Oliveira CS, Nunes BT, Cruz AC, Nascimento AG, Medeiros RC, Caldas CA, Araujo FC, Quaresma JA, Vasconcelos BC, Queiroz MG, da Rosa ES, Henriques DF, Silva EV, Chiang JO, Martins LC, Medeiros DB, Lima JA, Nunes MR, Cardoso JF, Silva SP, Shi PY, Tesh RB, Rodrigues SG, Vasconcelos PF. Zika virus epidemic in Brazil. I Fatal disease in adults: clinical and laboratorial aspects. J Clin Virol. 2016;85:56–64. - PMC - PubMed
-
- Barrows NJ, Campos RK, Powell ST, Prasanth KR, Schott-Lerner G, Soto-Acosta R, Galarza-Munoz G, McGrath EL, Urrabaz-Garza R, Gao J, Wu P, Menon R, Saade G, Fernandez-Salas I, Rossi SL, Vasilakis N, Routh A, Bradrick SS, Garcia-Blanco MA. A screen of FDA-approved drugs for inhibitors of Zika virus infection. Cell Host Microbe. 2016;20:259–270. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
