

Sirtuin 2 mutations in human cancers impair its function in genome maintenance
- PMID: 28461331
- PMCID: PMC5473244
- DOI: 10.1074/jbc.M116.772566
Sirtuin 2 mutations in human cancers impair its function in genome maintenance
Abstract
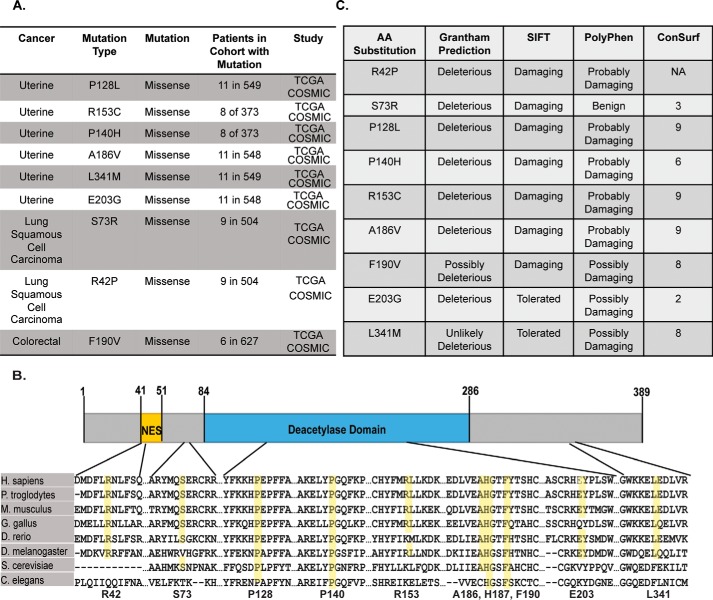
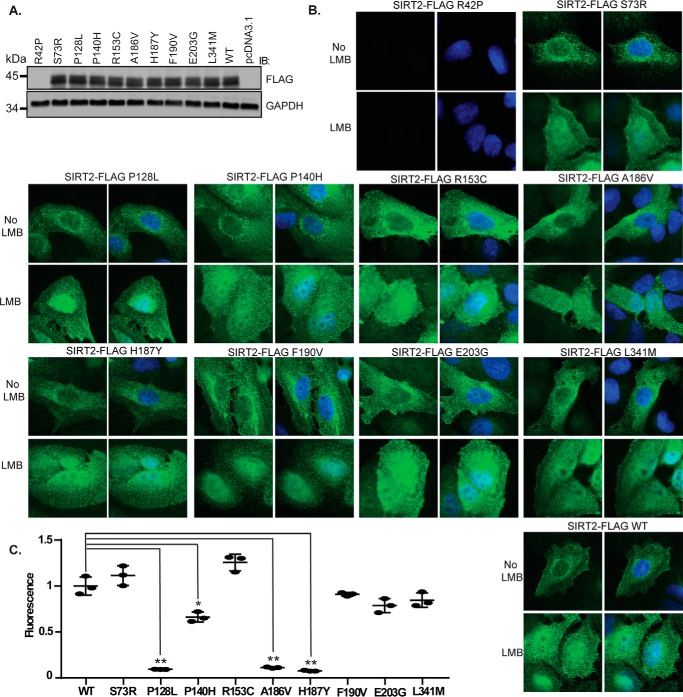
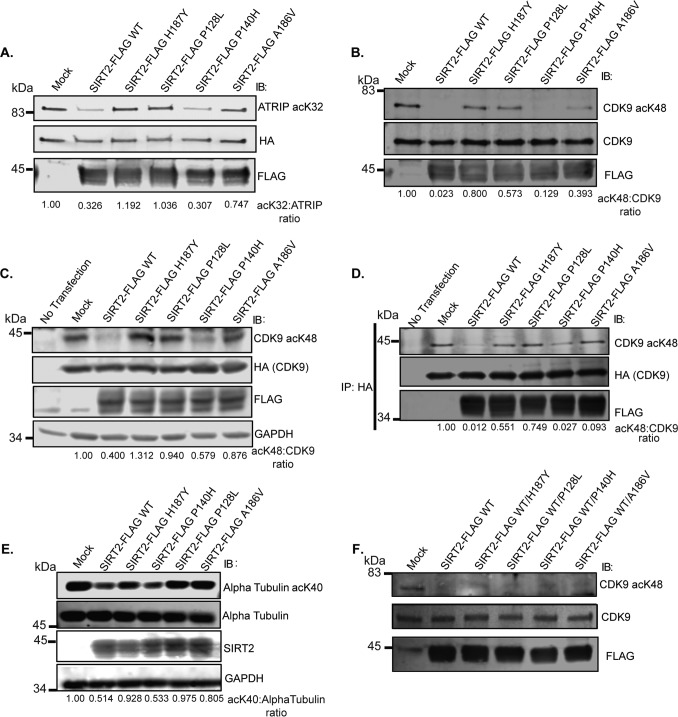
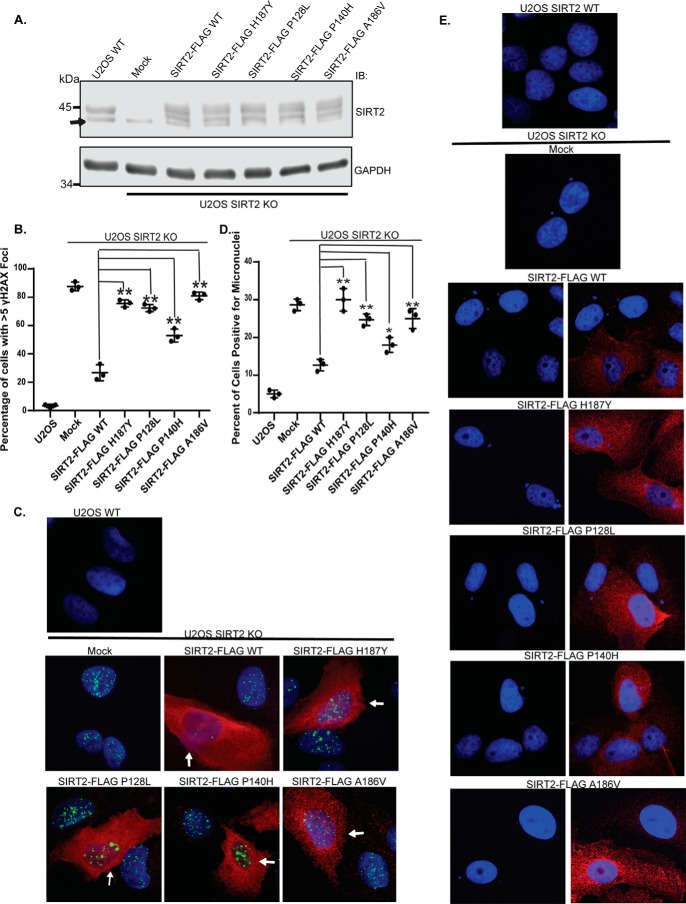
Sirtuin 2 (SIRT2) is a sirtuin family deacetylase, which maintains genome integrity and prevents tumorigenesis. Although Sirt2 deficiency in mice leads to tumorigenesis, the functional significance of somatic SIRT2 mutations in human tumors is unclear. Using structural insight combined with bioinformatics and functional analyses, we show that naturally occurring cancer-associated SIRT2 mutations at evolutionarily conserved sites disrupt its deacetylation of DNA-damage response proteins by impairing SIRT2 catalytic activity or protein levels but not its localization or binding with substrate. We observed that these SIRT2 mutant proteins fail to restore the replication stress sensitivity, impairment in recovery from replication stress, and impairment in ATR-interacting protein (ATRIP) focus accumulation of SIRT2 deficiency. Moreover, the SIRT2 mutant proteins failed to rescue the spontaneous induction of DNA damage and micronuclei of SIRT2 deficiency in cancer cells. Our findings support a model for SIRT2's tumor-suppressive function in which somatic mutations in SIRT2 contribute to genomic instability by impairing its deacetylase activity or diminishing its protein levels in the DNA-damage response. In conclusion, our work provides a mechanistic basis for understanding the biological and clinical significance of SIRT2 mutations in genome maintenance and tumor suppression.
Keywords: DNA; DNA damage; DNA damage response; acetylation; cancer; genomic instability; sirtuin; tumor suppressor gene.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures


Similar articles
-
ATRIP Deacetylation by SIRT2 Drives ATR Checkpoint Activation by Promoting Binding to RPA-ssDNA.Cell Rep. 2016 Feb 16;14(6):1435-1447. doi: 10.1016/j.celrep.2016.01.018. Epub 2016 Feb 4. Cell Rep. 2016. PMID: 26854234 Free PMC article.
-
SIRT2 directs the replication stress response through CDK9 deacetylation.Proc Natl Acad Sci U S A. 2013 Aug 13;110(33):13546-51. doi: 10.1073/pnas.1301463110. Epub 2013 Jul 29. Proc Natl Acad Sci U S A. 2013. PMID: 23898190 Free PMC article.
-
A novel sirtuin 2 (SIRT2) inhibitor with p53-dependent pro-apoptotic activity in non-small cell lung cancer.J Biol Chem. 2014 Feb 21;289(8):5208-16. doi: 10.1074/jbc.M113.487736. Epub 2013 Dec 30. J Biol Chem. 2014. PMID: 24379401 Free PMC article.
-
SIRT2: Controversy and multiple roles in disease and physiology.Ageing Res Rev. 2019 Nov;55:100961. doi: 10.1016/j.arr.2019.100961. Epub 2019 Sep 7. Ageing Res Rev. 2019. PMID: 31505260 Review.
-
The role of SIRT2 in cancer: A novel therapeutic target.Int J Cancer. 2020 Dec 15;147(12):3297-3304. doi: 10.1002/ijc.33118. Epub 2020 Jun 16. Int J Cancer. 2020. PMID: 32449165 Review.
Cited by
-
Fueling genome maintenance: On the versatile roles of NAD+ in preserving DNA integrity.J Biol Chem. 2022 Jun;298(6):102037. doi: 10.1016/j.jbc.2022.102037. Epub 2022 May 17. J Biol Chem. 2022. PMID: 35595095 Free PMC article. Review.
-
Sirtuins in B lymphocytes metabolism and function.World J Exp Med. 2019 Jan 16;9(1):1-13. doi: 10.5493/wjem.v9.i1.1. eCollection 2019 Jan 16. World J Exp Med. 2019. PMID: 30705866 Free PMC article. Review.
-
BubR1 and SIRT2: Insights into aneuploidy, aging, and cancer.Semin Cancer Biol. 2024 Nov;106-107:201-216. doi: 10.1016/j.semcancer.2024.10.005. Epub 2024 Oct 28. Semin Cancer Biol. 2024. PMID: 39490401 Review.
-
DNA Repair: Translation to the Clinic.Clin Oncol (R Coll Radiol). 2019 May;31(5):303-310. doi: 10.1016/j.clon.2019.02.007. Epub 2019 Mar 12. Clin Oncol (R Coll Radiol). 2019. PMID: 30876709 Free PMC article. Review.
-
Why Is Longevity Still a Scientific Mystery? Sirtuins-Past, Present and Future.Int J Mol Sci. 2022 Dec 31;24(1):728. doi: 10.3390/ijms24010728. Int J Mol Sci. 2022. PMID: 36614171 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
