

Mechanism of ubiquitin chain synthesis employed by a HECT domain ubiquitin ligase
- PMID: 28461335
- PMCID: PMC5481553
- DOI: 10.1074/jbc.M117.789479
Mechanism of ubiquitin chain synthesis employed by a HECT domain ubiquitin ligase
Abstract
Keywords: E3 ubiquitin ligase; HECT; polyubiquitin chain; protein degradation; ubiquitin; ubiquitylation (ubiquitination).
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
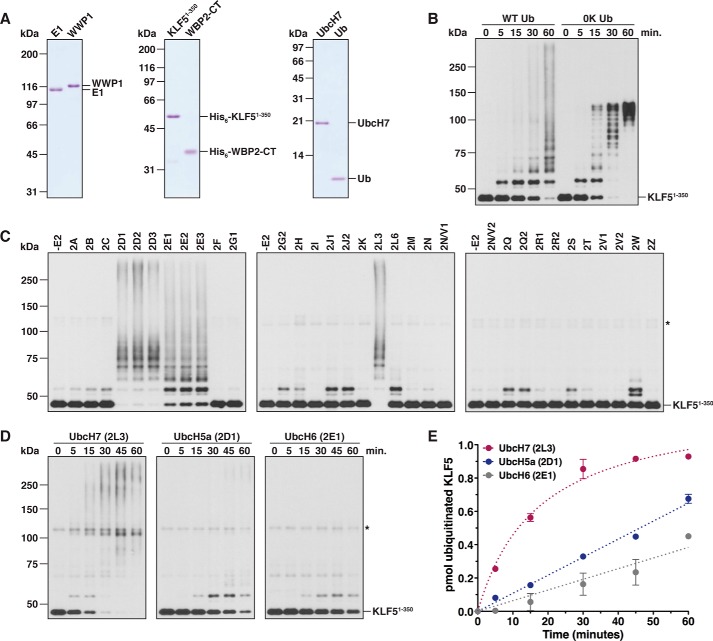
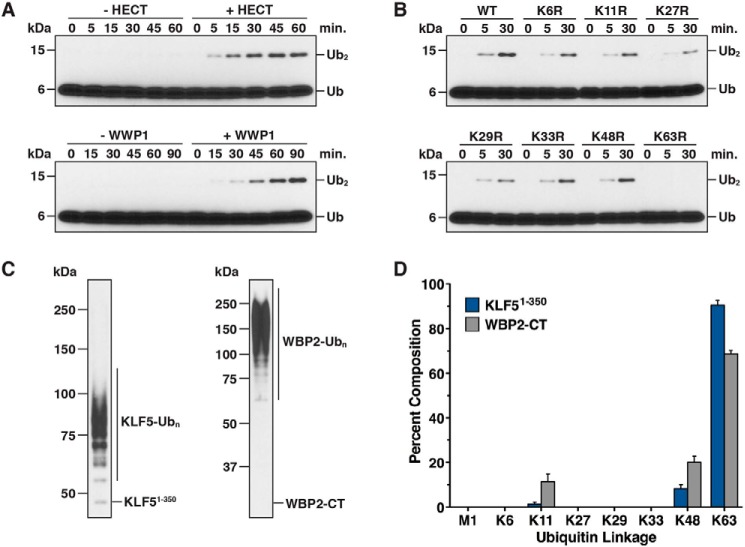
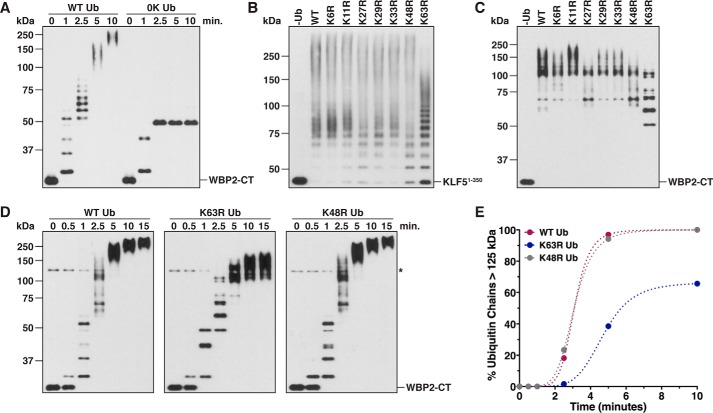
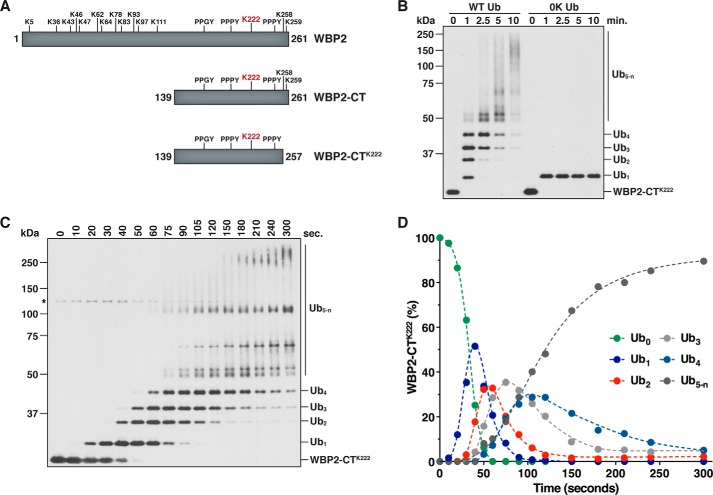
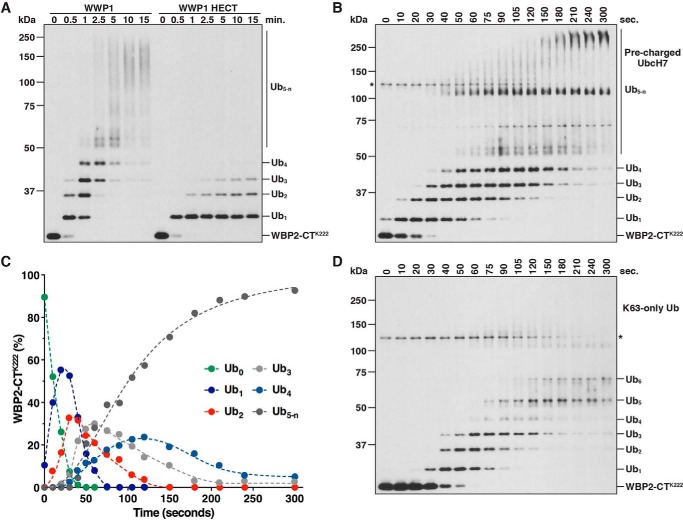
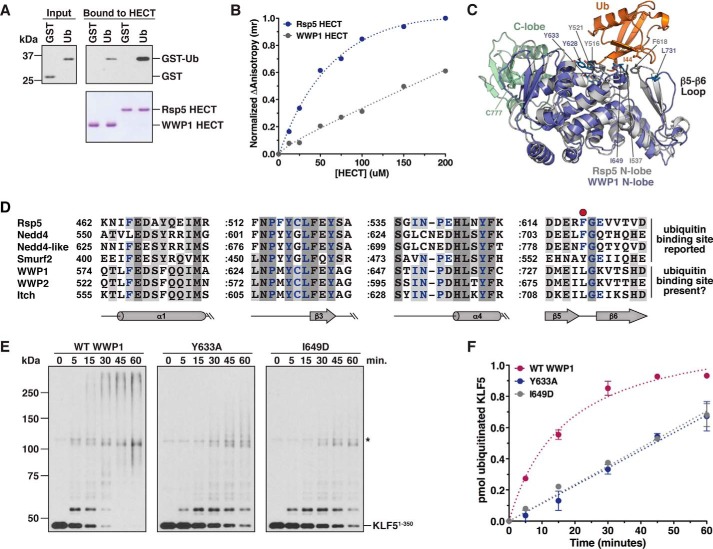
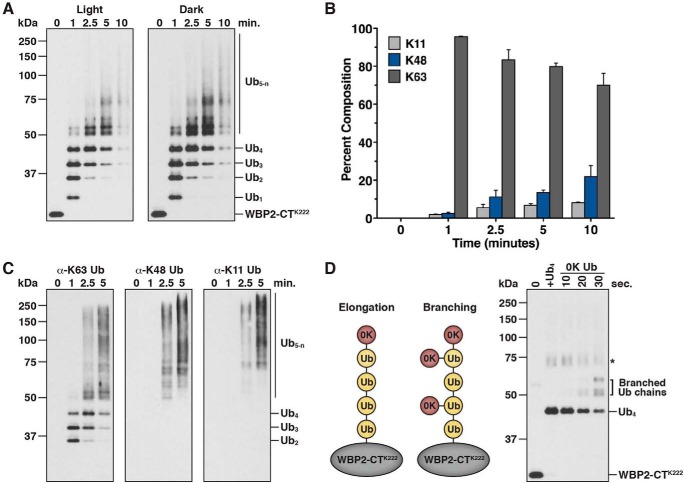
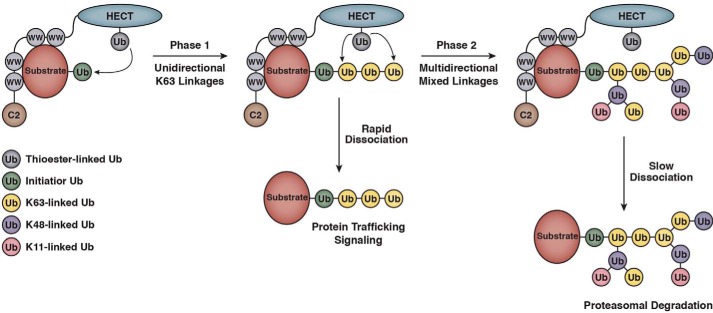
References
-
- Komander D., and Rape M. (2012) The ubiquitin code. Annu. Rev. Biochem. 81, 203–229 - PubMed
-
- Kulathu Y., and Komander D. (2012) Atypical ubiquitylation–the unexplored world of polyubiquitin beyond Lys48 and Lys63 linkages. Nat. Rev. Mol. Cell Biol. 13, 508–523 - PubMed
-
- Husnjak K., and Dikic I. (2012) Ubiquitin-binding proteins: decoders of ubiquitin-mediated cellular functions. Annu. Rev. Biochem. 81, 291–322 - PubMed
-
- Deshaies R. J., and Joazeiro C. A. (2009) RING domain E3 ubiquitin ligases. Annu. Rev. Biochem. 78, 399–434 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
