

SP and IL-33 together markedly enhance TNF synthesis and secretion from human mast cells mediated by the interaction of their receptors
- PMID: 28461492
- PMCID: PMC5441798
- DOI: 10.1073/pnas.1524845114
SP and IL-33 together markedly enhance TNF synthesis and secretion from human mast cells mediated by the interaction of their receptors
Abstract
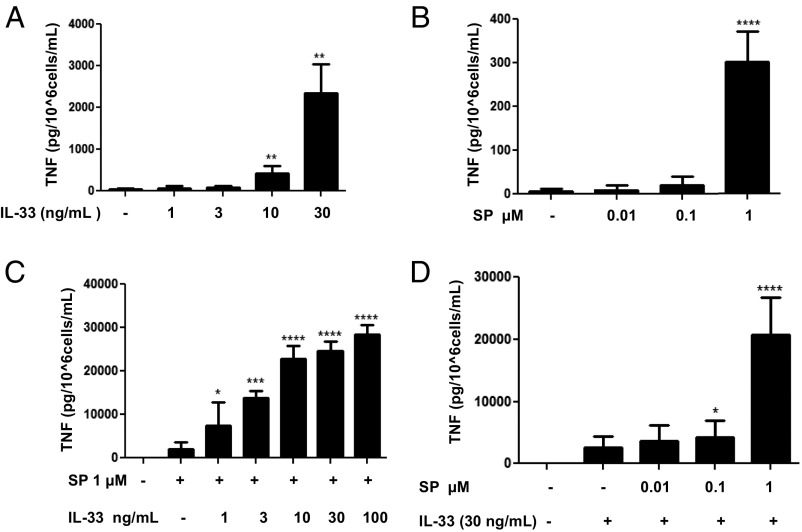
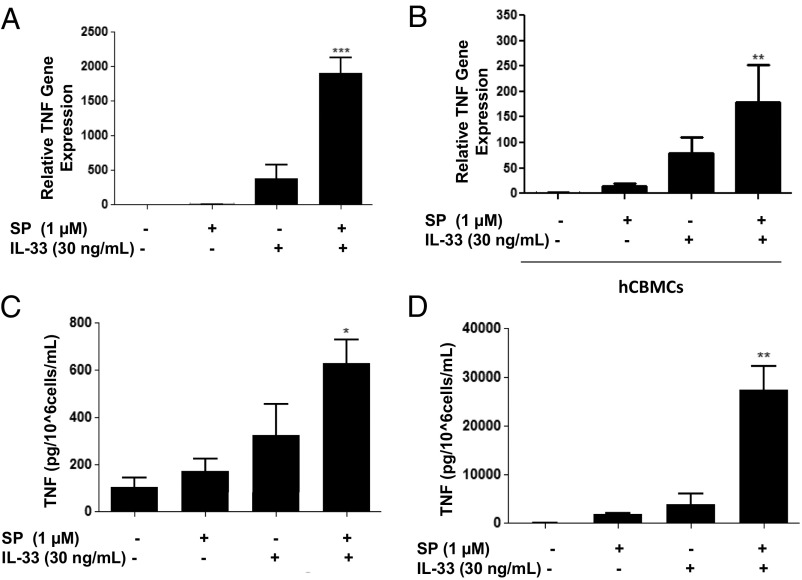
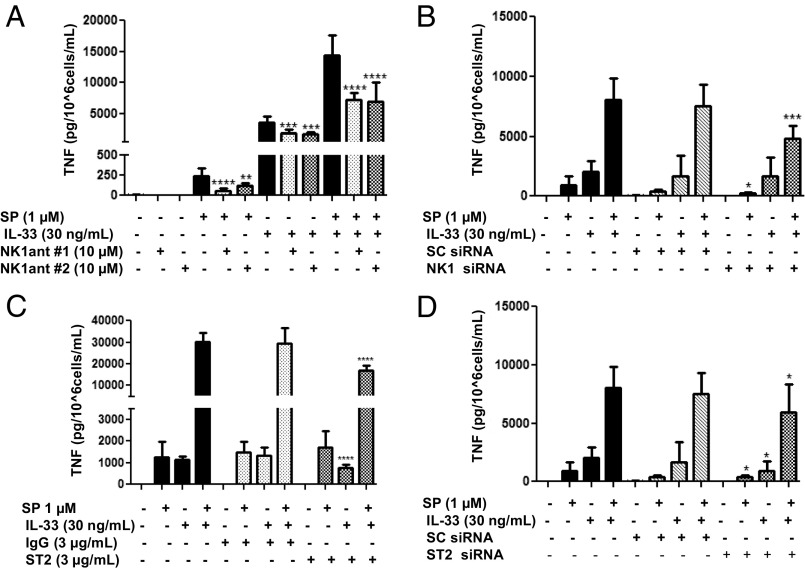
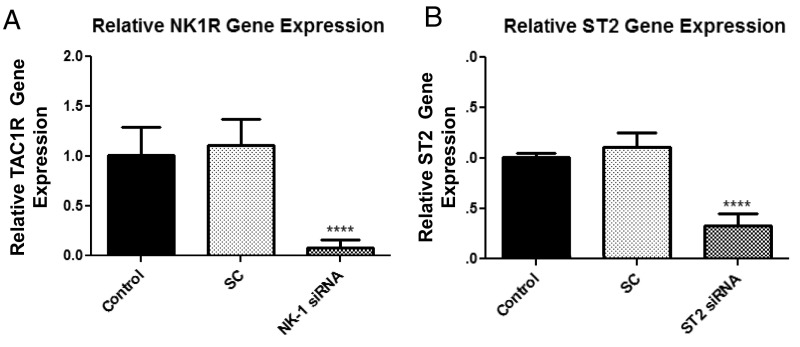
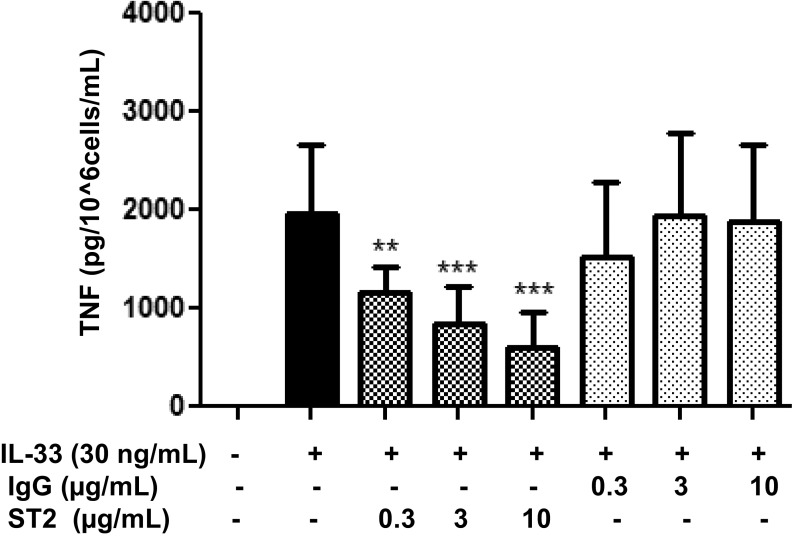
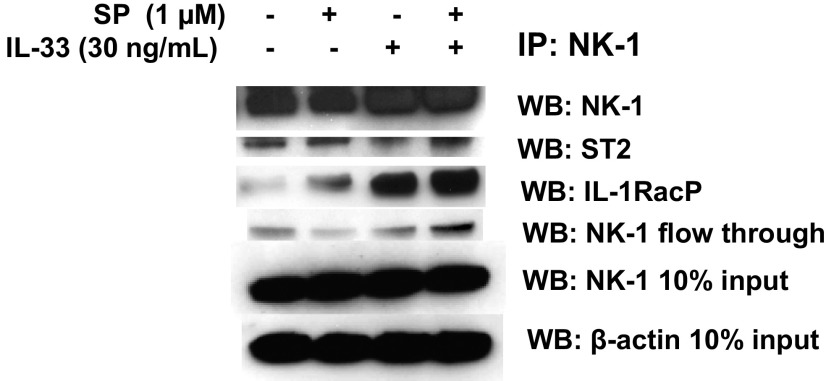
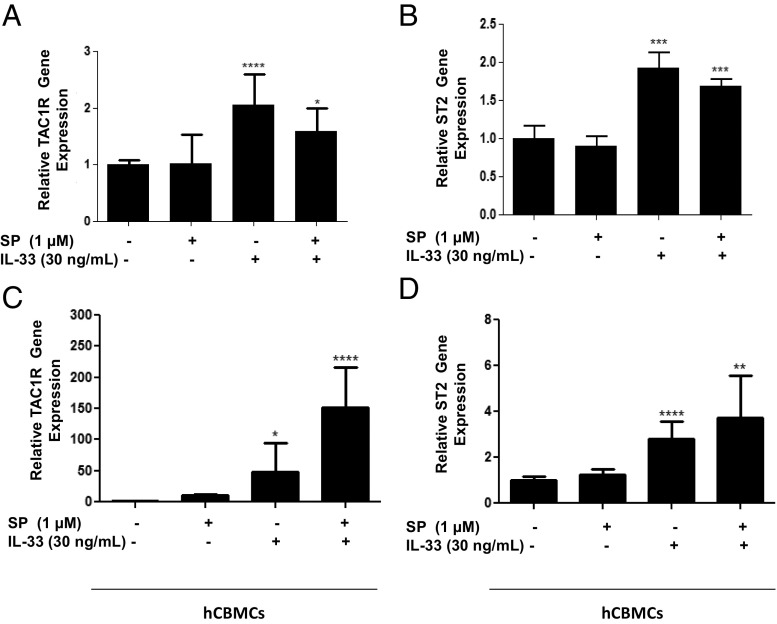
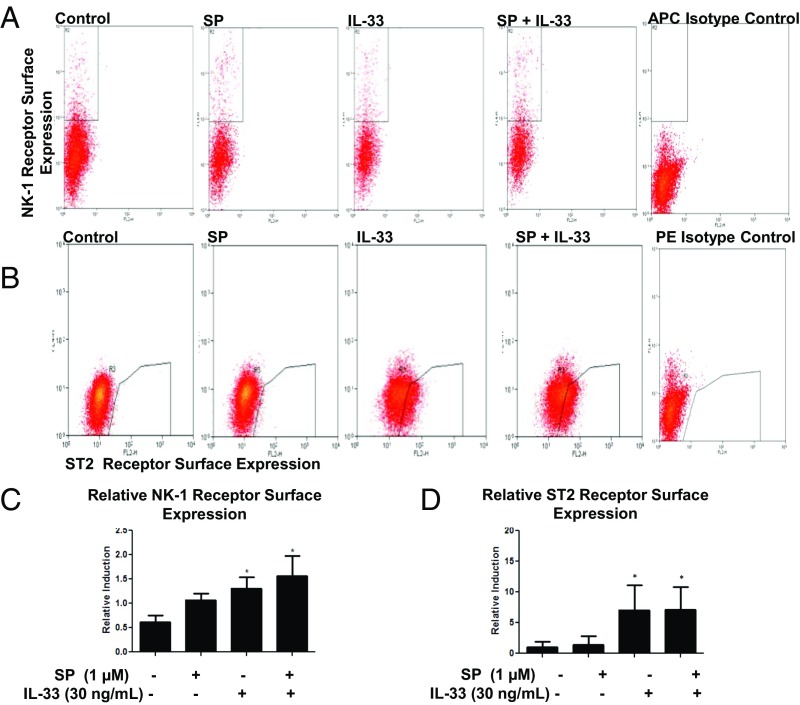
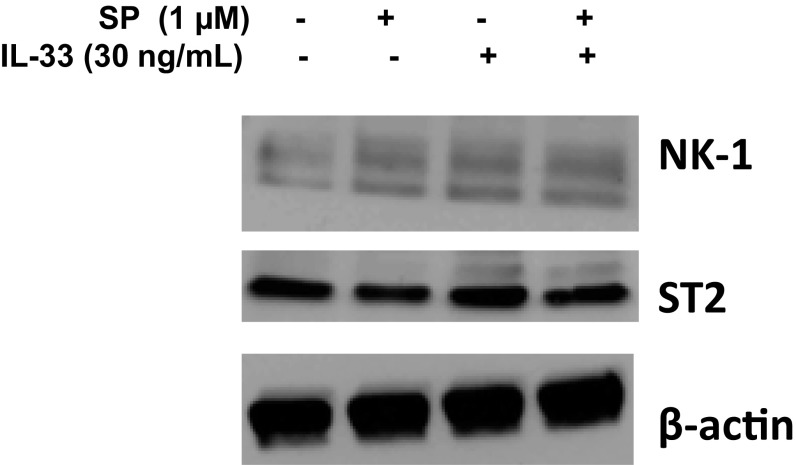
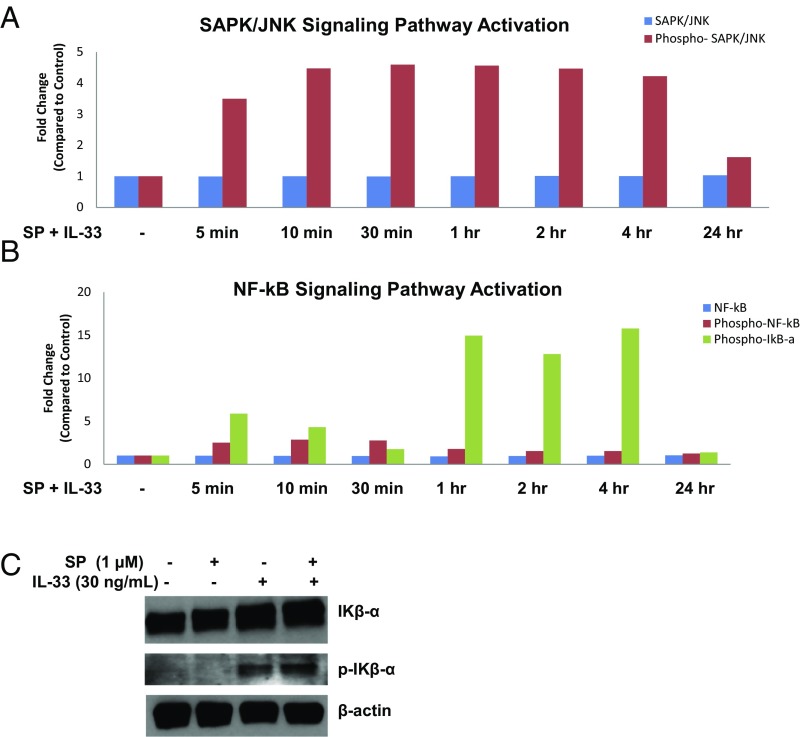
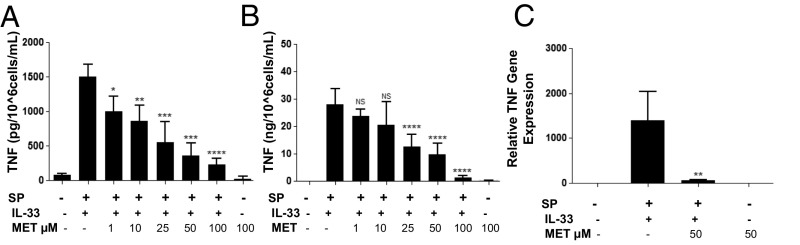
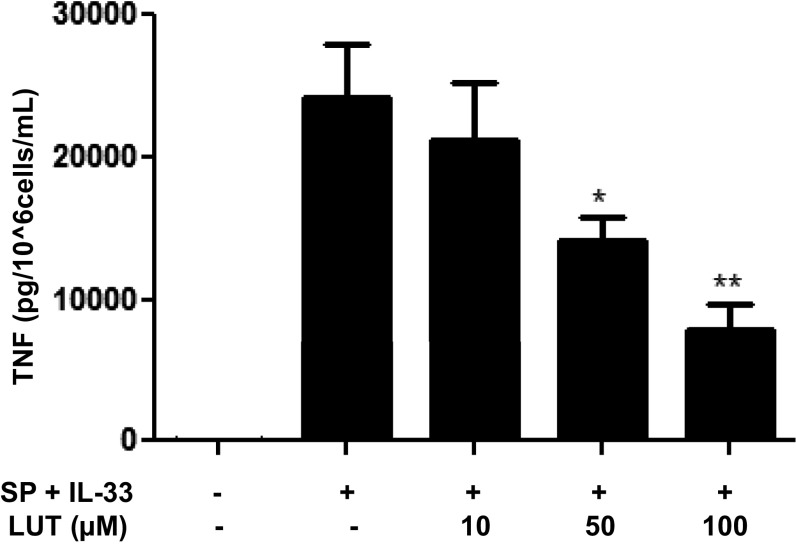
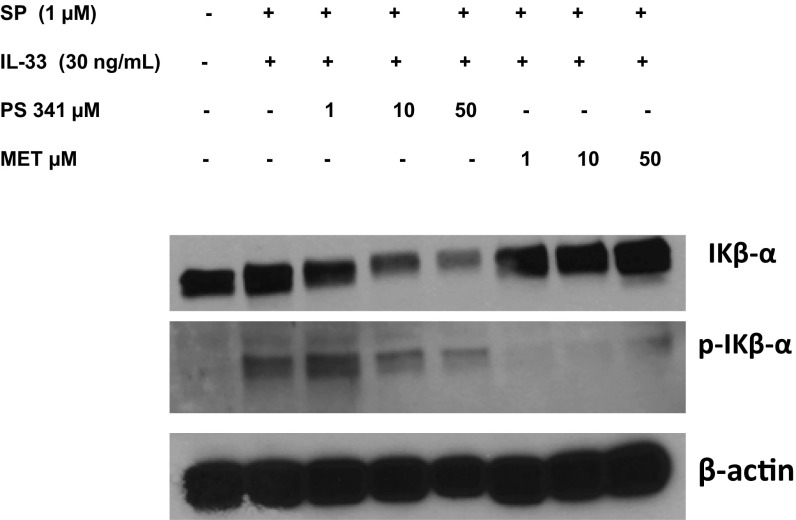
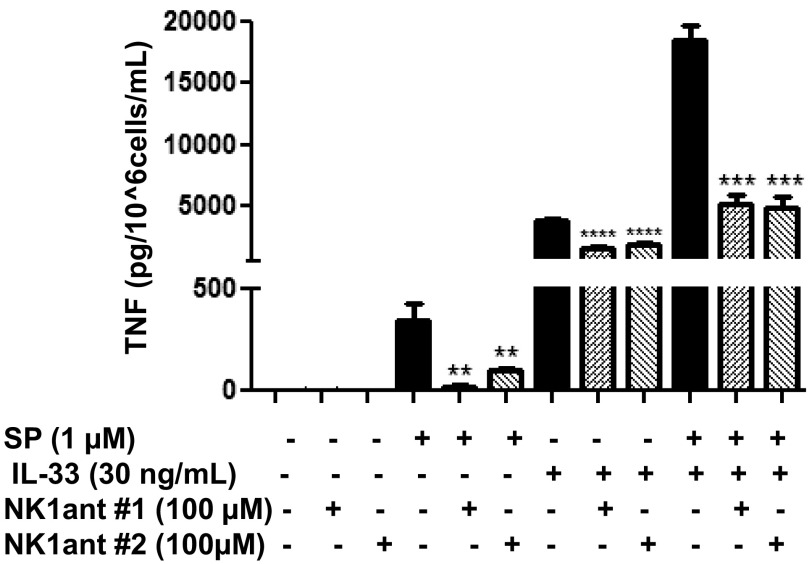
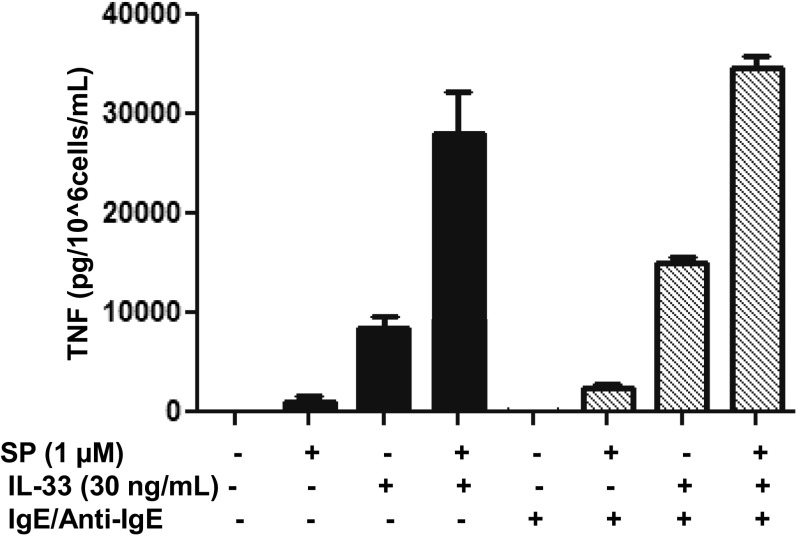
The peptide substance P (SP) and the cytokine tumor necrosis factor (TNF) have been implicated in inflammatory processes. Mast cells are recognized as important in inflammatory responses. Here, we report that IL-33 (30 ng/mL), a member of the IL-1 family of cytokines, administered in combination with SP (1 µM), markedly increase (by 1,000-fold) TNF gene expression in cultured human LAD2 and primary mast cells derived from umbilical cord blood. SP (0.01-1 μM) and IL-33 (1-100 ng/mL) in combination also greatly stimulate TNF secretion (by 4,500-fold). Pretreatment of LAD2 cells with two different neurokinin-1 (NK-1) receptor antagonists and siRNA inhibits TNF secretion by 50% (P < 0.001) when stimulated by SP and IL-33. Pretreatment of LAD2 cells with a neutralizing antibody for IL-33 receptor, ST2, inhibits TNF secretion by 50% (P < 0.001), and ST2 siRNA decreases TNF secretion by 30% (P < 0.05), when stimulated by SP and IL-33. Surprisingly, NK-1 antagonists also inhibit 50% of TNF secretion (P < 0.001) when stimulated only by IL-33, and ST2 receptor reduction also decreases SP-stimulated TNF secretion by 30% (P < 0.05), suggesting an interaction between NK-1 and ST2 receptors. Moreover, IL-33 increases NK-1 gene and surface protein expression, as well as IKβ-α phosphorylation. Pretreatment of LAD2 cells with 5,7,3',4'-tetramethoxyflavone (methoxyluteolin) (1-100 μM) inhibits (P < 0.001) TNF gene expression (98%) and secretion (64%) at 50 µM and phosphorylation of p-IKB-α at 1 μM when stimulated by SP and IL-33. These findings identify a unique amplification process of TNF synthesis and secretion via the interaction of NK-1 and ST2 receptors inhibitable by methoxyluteolin.
Keywords: inflammation; interleukin-33; mast cells; substance P; tumor necrosis factor.
Conflict of interest statement
Conflict of interest statement: T.C.T. and Cem Akin were coauthors on a 2015 publication. This publication was a review article and did not involve any research collaboration.
Figures

Comment in
-
Reply to Fattori et al.: Action of SP and IL-33 on mast cells.Proc Natl Acad Sci U S A. 2017 Nov 21;114(47):E10036. doi: 10.1073/pnas.1716233114. Epub 2017 Nov 6. Proc Natl Acad Sci U S A. 2017. PMID: 29109296 Free PMC article. No abstract available.
-
IL-33/ST2 signaling boosts inflammation and pain.Proc Natl Acad Sci U S A. 2017 Nov 21;114(47):E10034-E10035. doi: 10.1073/pnas.1716120114. Epub 2017 Nov 6. Proc Natl Acad Sci U S A. 2017. PMID: 29109297 Free PMC article. No abstract available.
References
-
- Chang MM, Leeman SE. Isolation of a sialogogic peptide from bovine hypothalamic tissue and its characterization as substance P. J Biol Chem. 1970;245:4784–4790. - PubMed
-
- Saraceno R, Kleyn CE, Terenghi G, Griffiths CE. The role of neuropeptides in psoriasis. Br J Dermatol. 2006;155:876–882. - PubMed
-
- O’Connor TM, et al. The role of substance P in inflammatory disease. J Cell Physiol. 2004;201:167–180. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
