

Long-term self-renewing human epicardial cells generated from pluripotent stem cells under defined xeno-free conditions
- PMID: 28462012
- PMCID: PMC5408455
- DOI: 10.1038/s41551-016-0003
Long-term self-renewing human epicardial cells generated from pluripotent stem cells under defined xeno-free conditions
Abstract
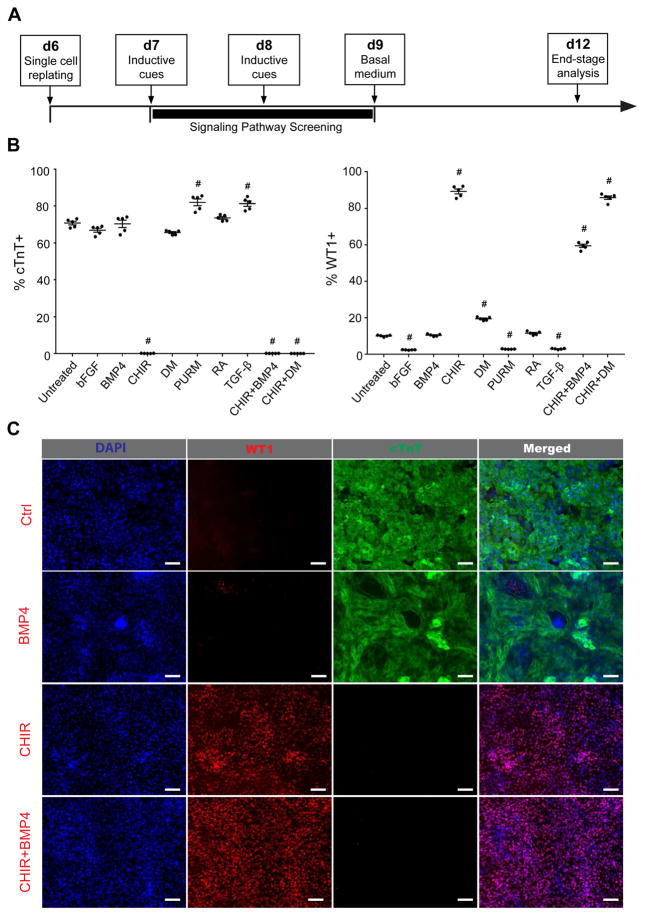
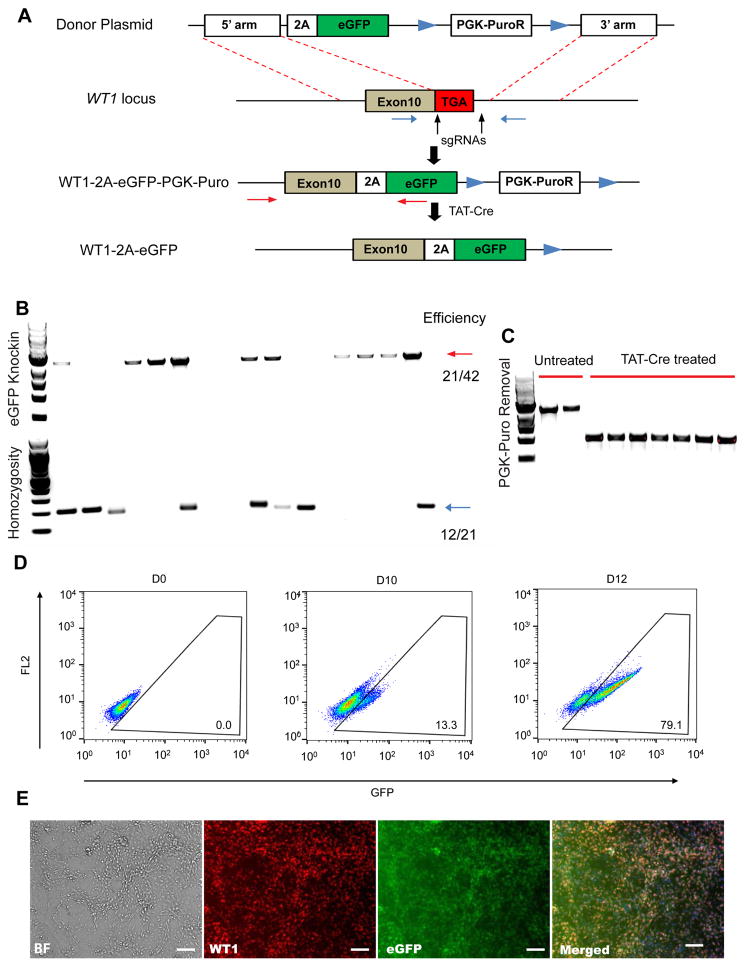
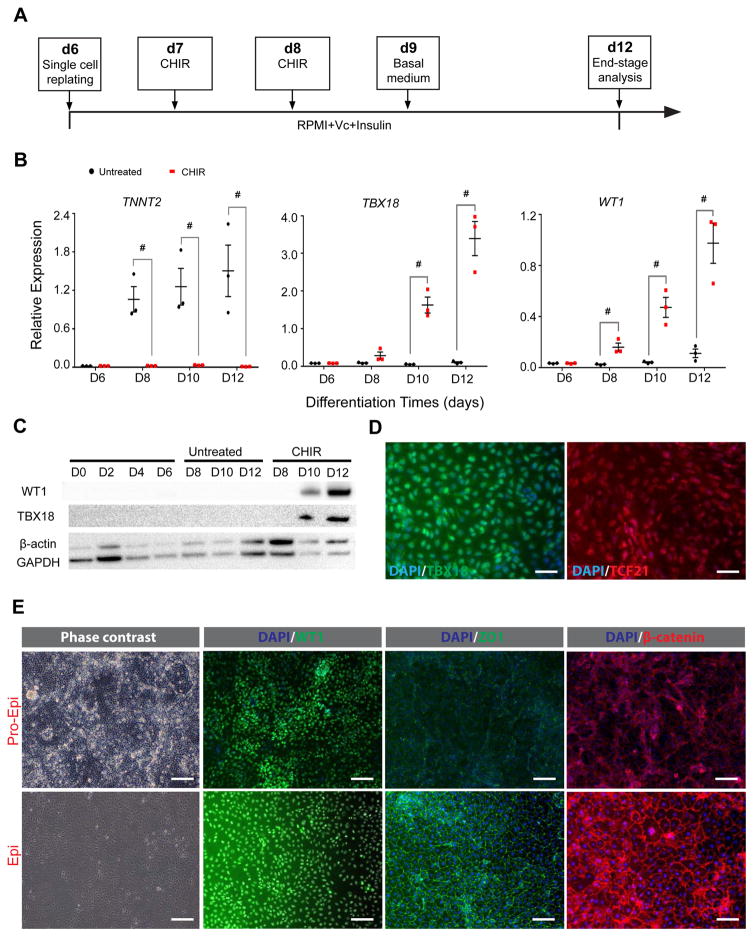
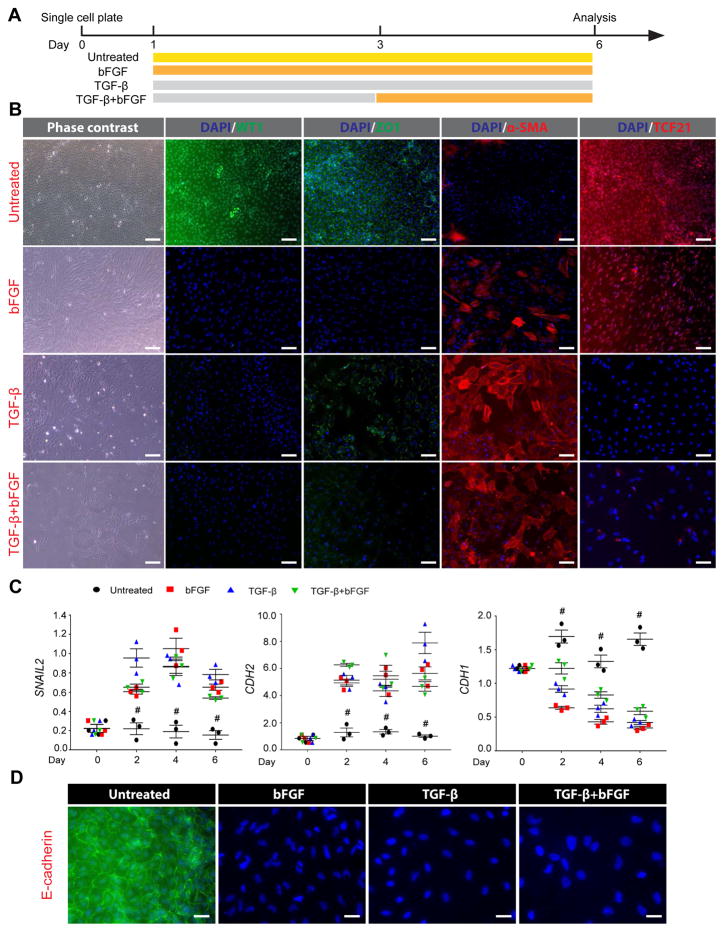
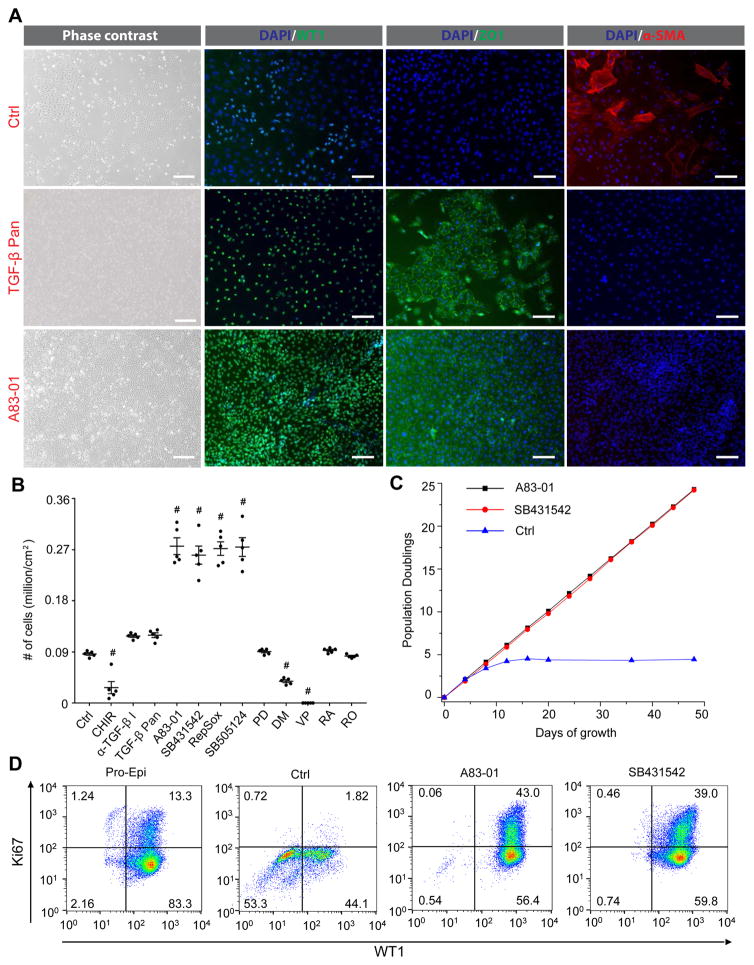
The epicardium contributes both multi-lineage descendants and paracrine factors to the heart during cardiogenesis and cardiac repair, underscoring its potential for cardiac regenerative medicine. Yet little is known about the cellular and molecular mechanisms that regulate human epicardial development and regeneration. Here, we show that the temporal modulation of canonical Wnt signaling is sufficient for epicardial induction from 6 different human pluripotent stem cell (hPSC) lines, including a WT1-2A-eGFP knock-in reporter line, under chemically-defined, xeno-free conditions. We also show that treatment with transforming growth factor beta (TGF-β)-signalling inhibitors permitted long-term expansion of the hPSC-derived epicardial cells, resulting in a more than 25 population doublings of WT1+ cells in homogenous monolayers. The hPSC-derived epicardial cells were similar to primary epicardial cells both in vitro and in vivo, as determined by morphological and functional assays, including RNA-seq. Our findings have implications for the understanding of self-renewal mechanisms of the epicardium and for epicardial regeneration using cellular or small-molecule therapies.
Conflict of interest statement
COMPETING FINANCIAL INTERESTS The authors declare no competing financial interests.
Figures

References
-
- Männer J, Ruiz-Lozano P. Development and Function of the Epicardium. Adv Dev Biol. 2007;18:333–357.
-
- Pérez-Pomares JM, et al. Origin of coronary endothelial cells from epicardial mesothelium in avian embryos. Int J Dev Biol. 2002;46:1005–13. - PubMed
-
- Smart N, et al. Thymosin beta4 induces adult epicardial progenitor mobilization and neovascularization. Nature. 2007;445:177–82. - PubMed
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials