

CD1b-autoreactive T cells contribute to hyperlipidemia-induced skin inflammation in mice
- PMID: 28463230
- PMCID: PMC5451229
- DOI: 10.1172/JCI92217
CD1b-autoreactive T cells contribute to hyperlipidemia-induced skin inflammation in mice
Abstract
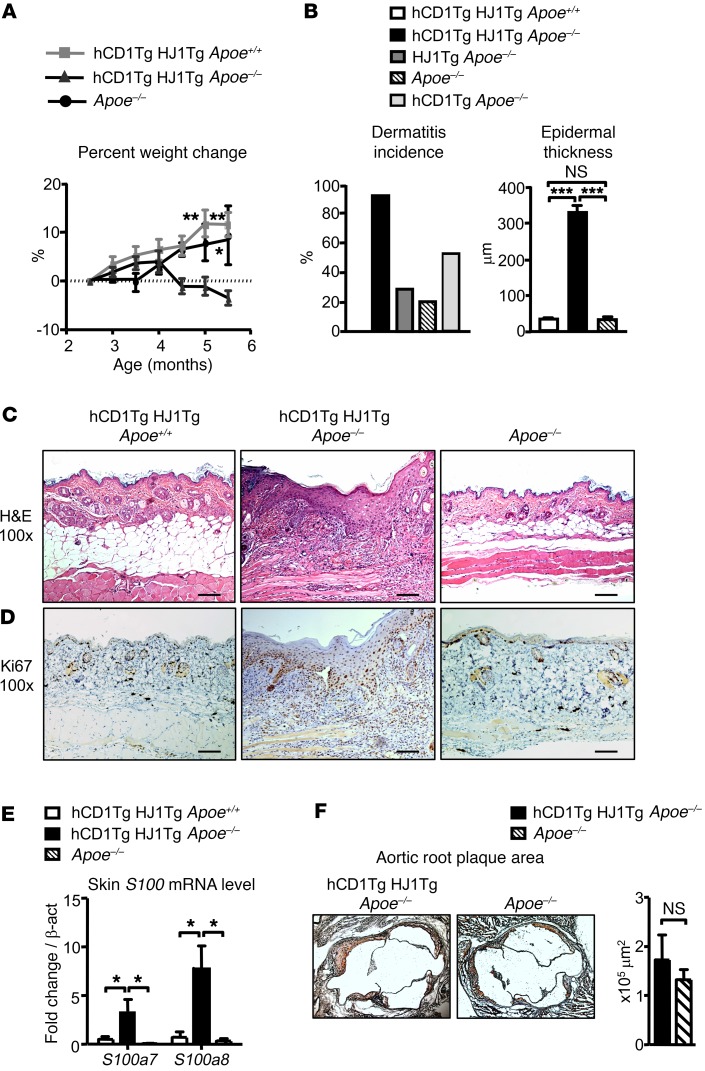
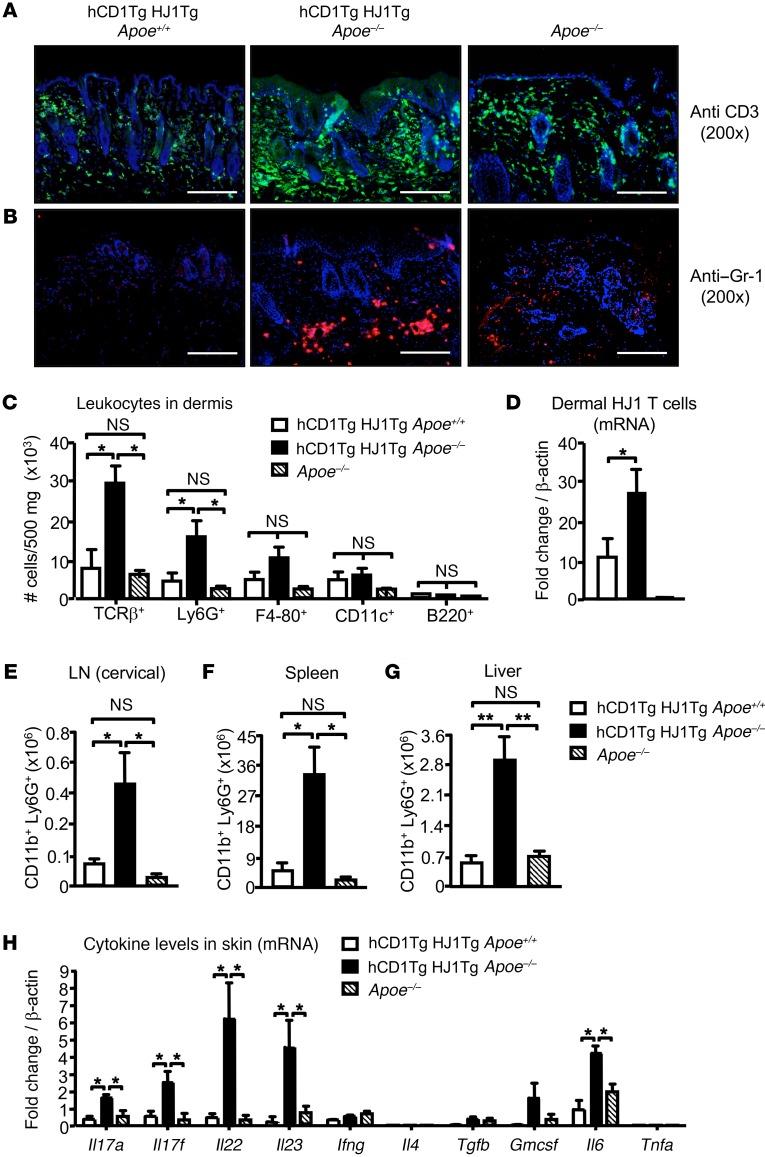
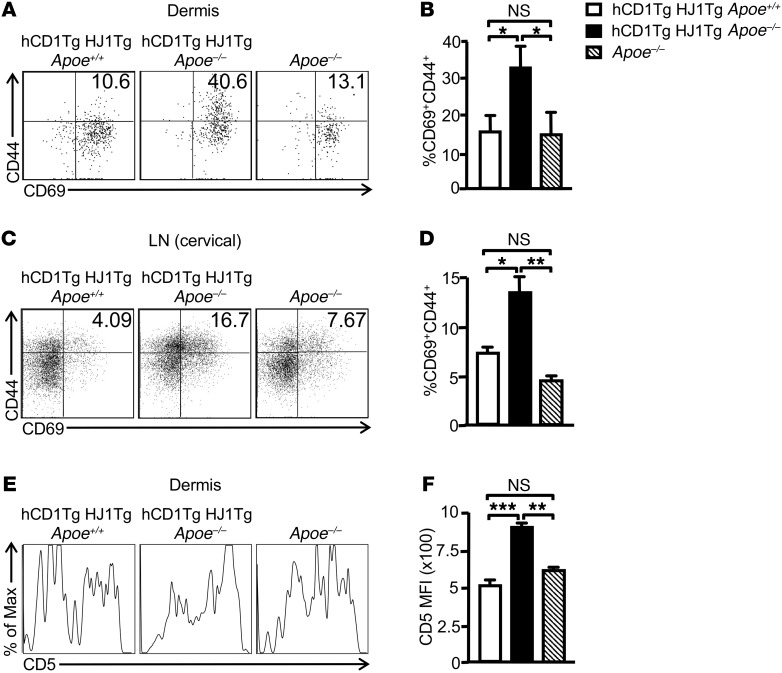
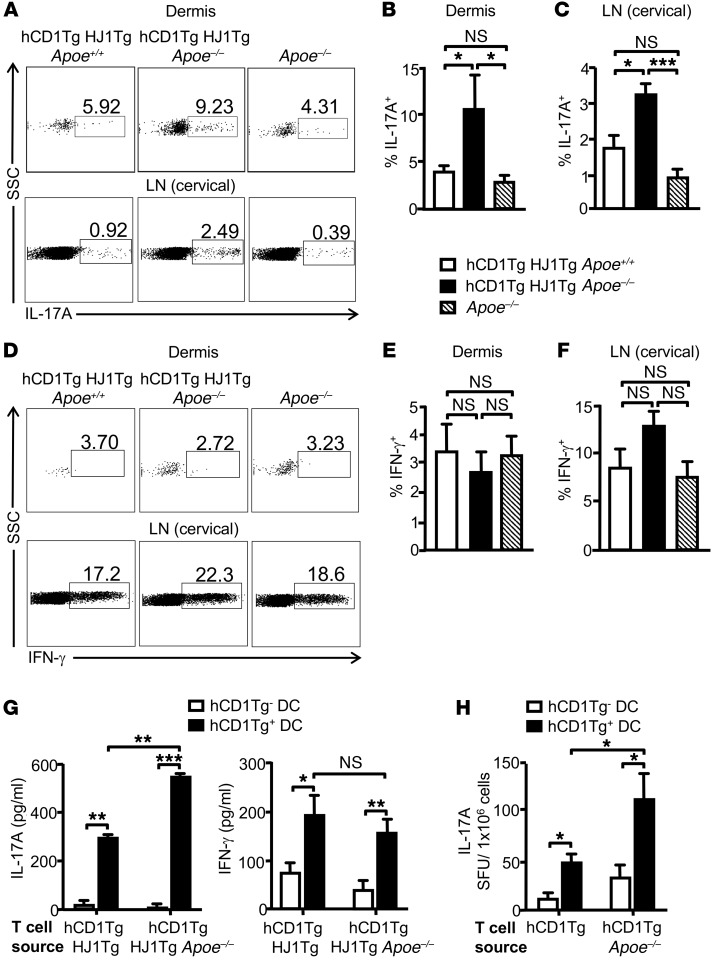
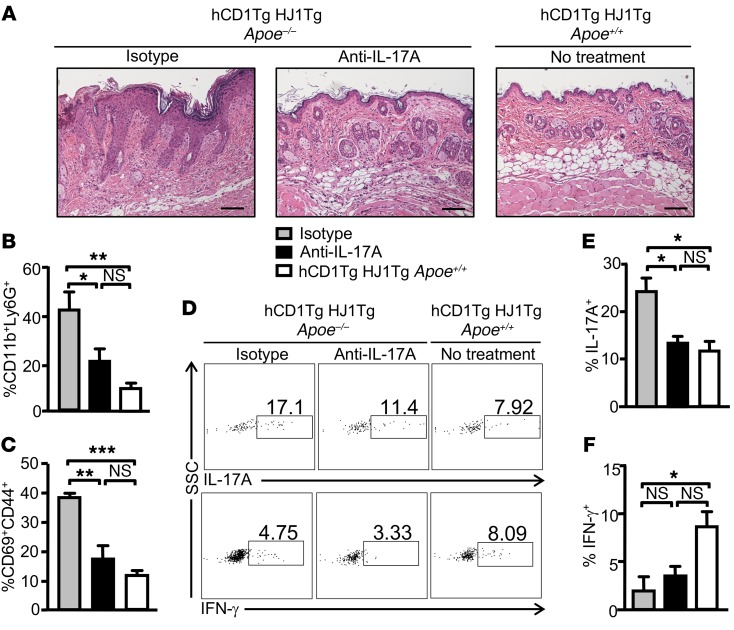
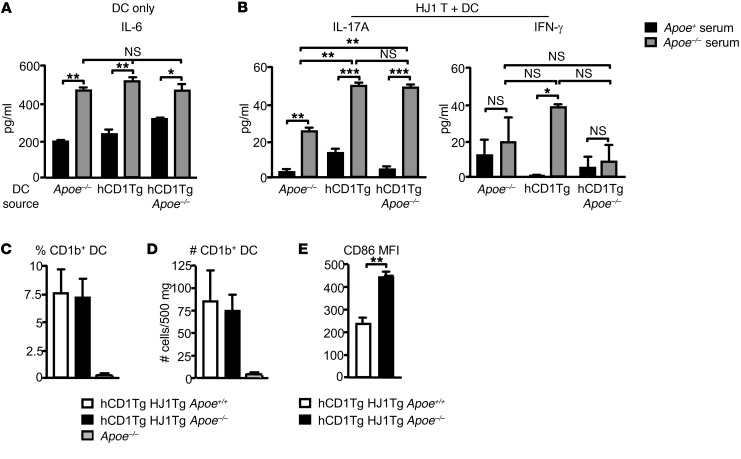
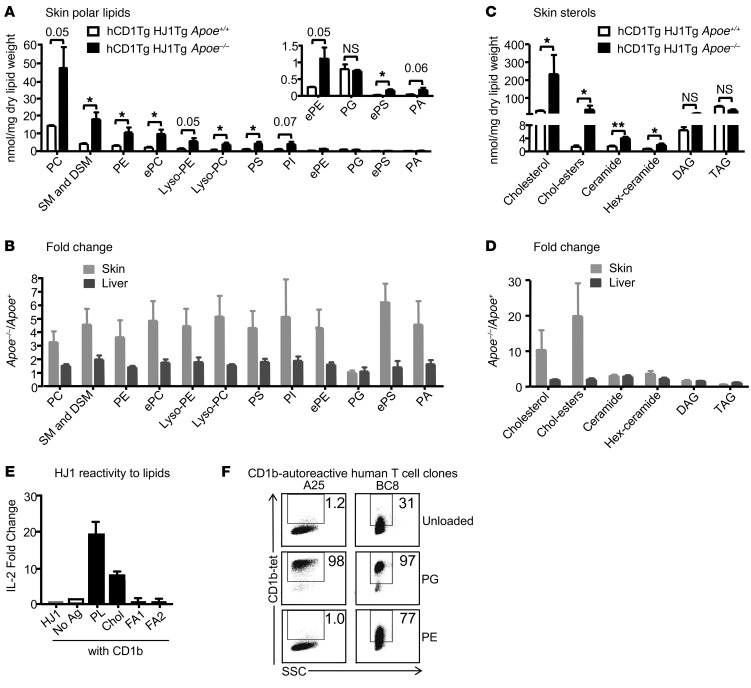
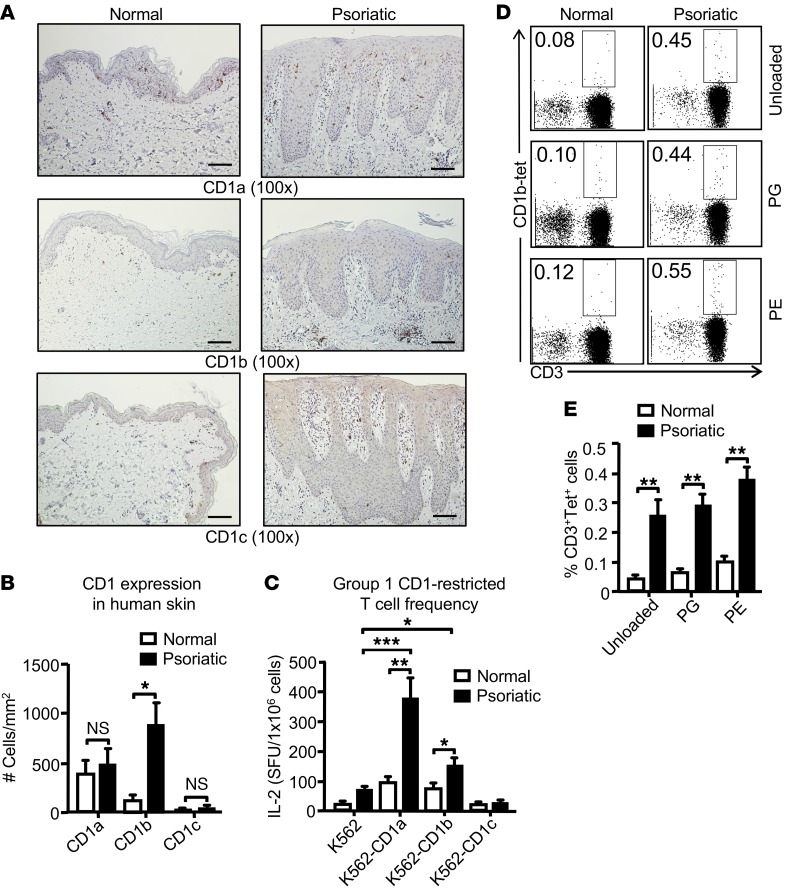
A large proportion of human T cells are autoreactive to group 1 CD1 proteins, which include CD1a, CD1b, and CD1c. However, the physiological role of the CD1 proteins remains poorly defined. Here, we have generated a double-transgenic mouse model that expresses human CD1b and CD1c molecules (hCD1Tg) as well as a CD1b-autoreactive TCR (HJ1Tg) in the ApoE-deficient background (hCD1Tg HJ1Tg Apoe-/- mice) to determine the role of CD1-autoreactive T cells in hyperlipidemia-associated inflammatory diseases. We found that hCD1Tg HJ1Tg Apoe-/- mice spontaneously developed psoriasiform skin inflammation characterized by T cell and neutrophil infiltration and a Th17-biased cytokine response. Anti-IL-17A treatment ameliorated skin inflammation in vivo. Additionally, phospholipids and cholesterol preferentially accumulated in diseased skin and these autoantigens directly activated CD1b-autoreactive HJ1 T cells. Furthermore, hyperlipidemic serum enhanced IL-6 secretion by CD1b+ DCs and increased IL-17A production by HJ1 T cells. In psoriatic patients, the frequency of CD1b-autoreactive T cells was increased compared with that in healthy controls. Thus, this study has demonstrated the pathogenic role of CD1b-autoreactive T cells under hyperlipidemic conditions in a mouse model of spontaneous skin inflammation. As a large proportion of psoriatic patients are dyslipidemic, this finding is of clinical significance and indicates that self-lipid-reactive T cells might serve as a possible link between hyperlipidemia and psoriasis.
Conflict of interest statement
Figures
References
-
- Ogden CL, Carroll MD, Fryar CD, Flegal KM. Prevalence of obesity among adults youth: United States, 2011-2014. NCHS Data Brief. 2015;(219):1–8. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
