

Cystinosin, the small GTPase Rab11, and the Rab7 effector RILP regulate intracellular trafficking of the chaperone-mediated autophagy receptor LAMP2A
- PMID: 28465352
- PMCID: PMC5481548
- DOI: 10.1074/jbc.M116.764076
Cystinosin, the small GTPase Rab11, and the Rab7 effector RILP regulate intracellular trafficking of the chaperone-mediated autophagy receptor LAMP2A
Abstract
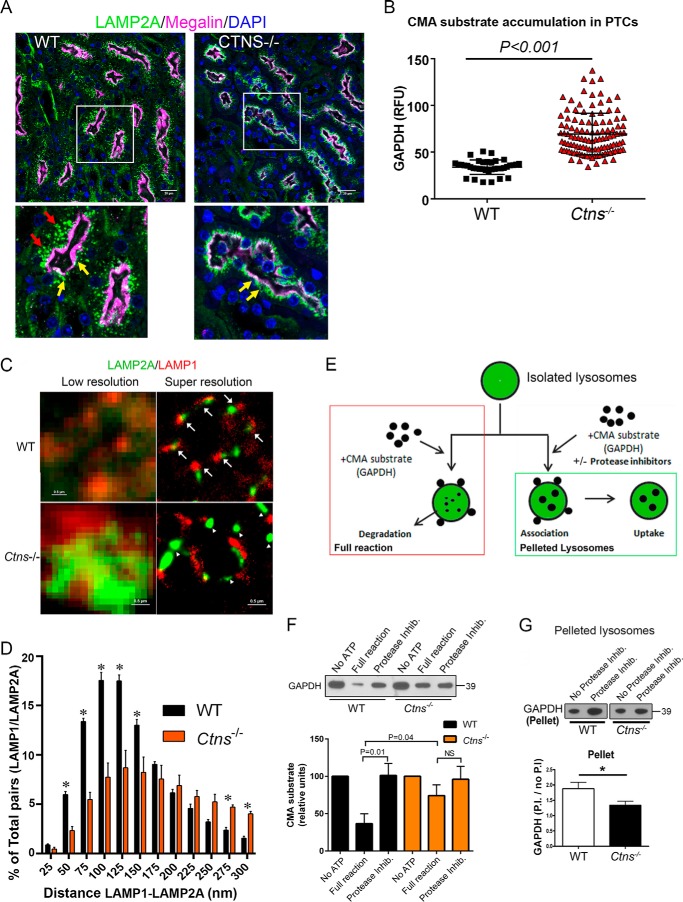
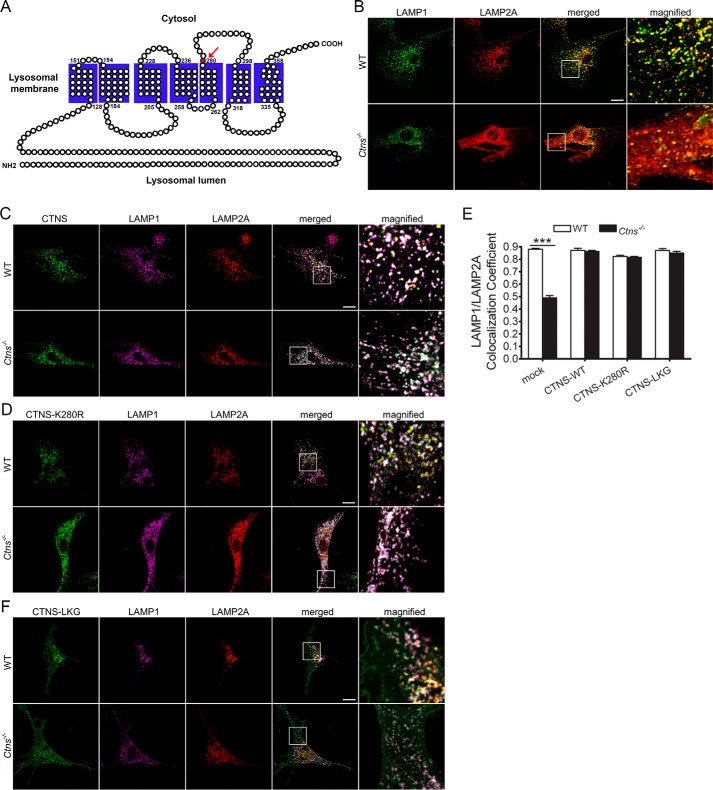
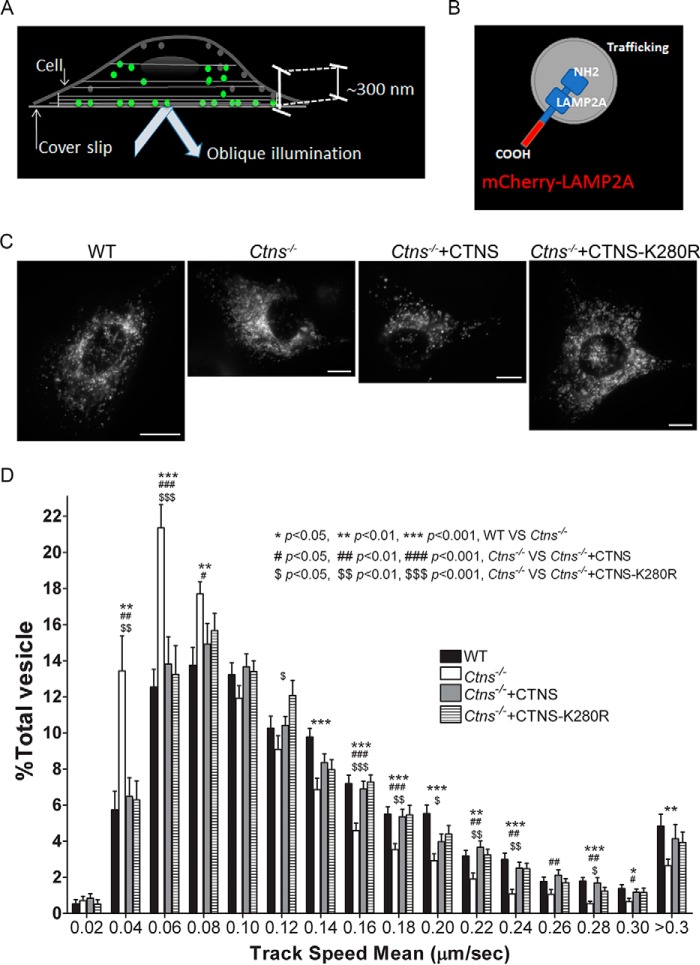
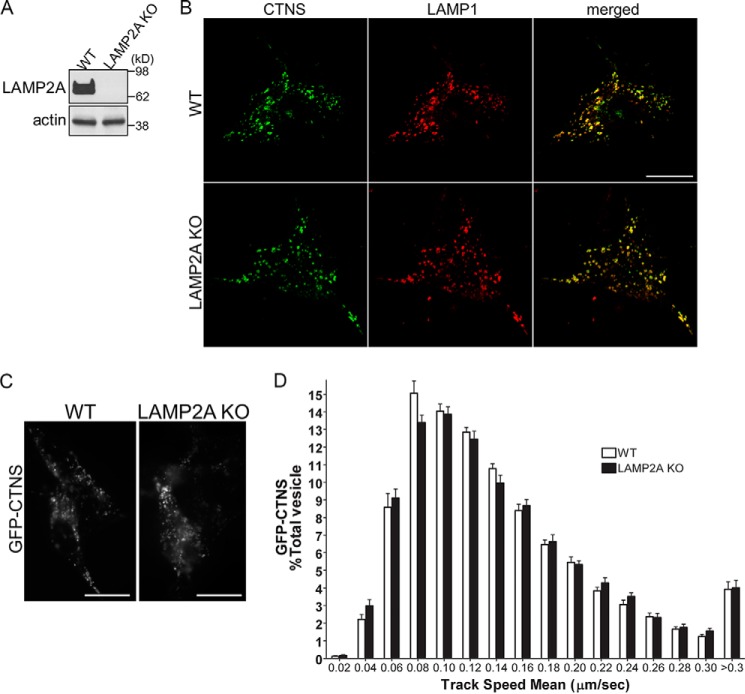
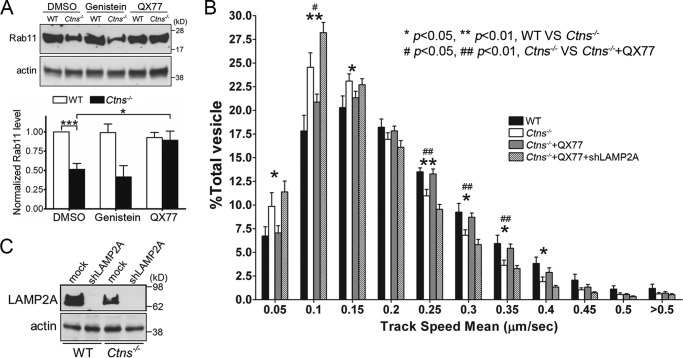
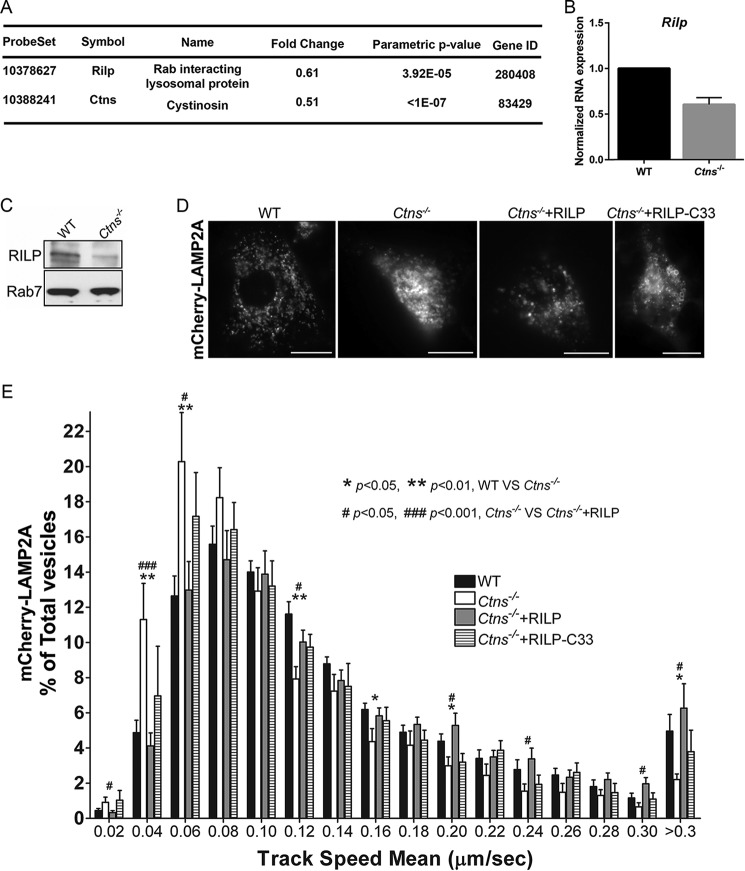
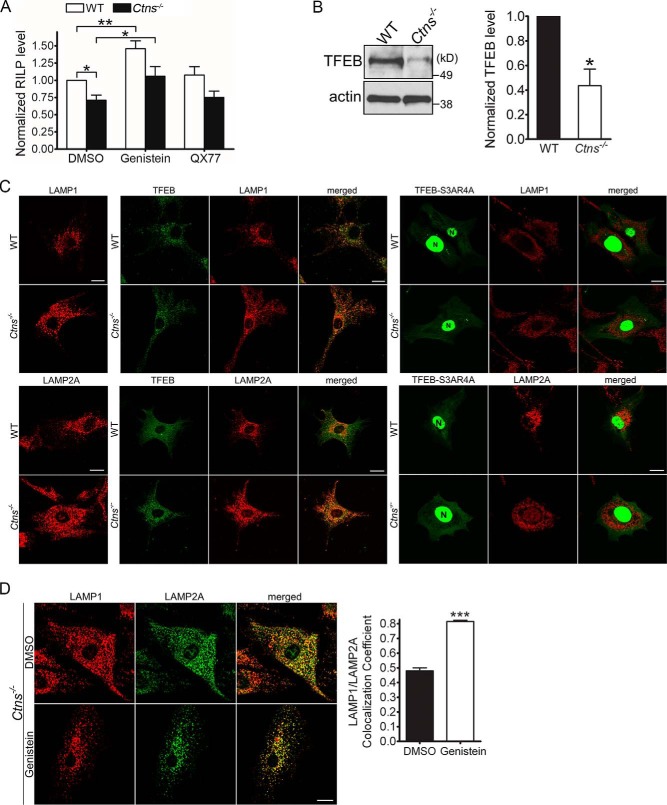
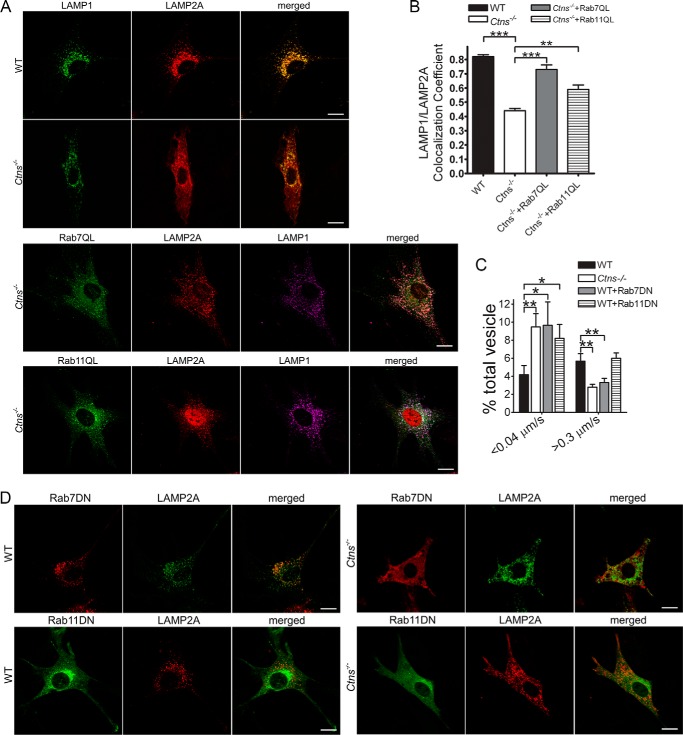
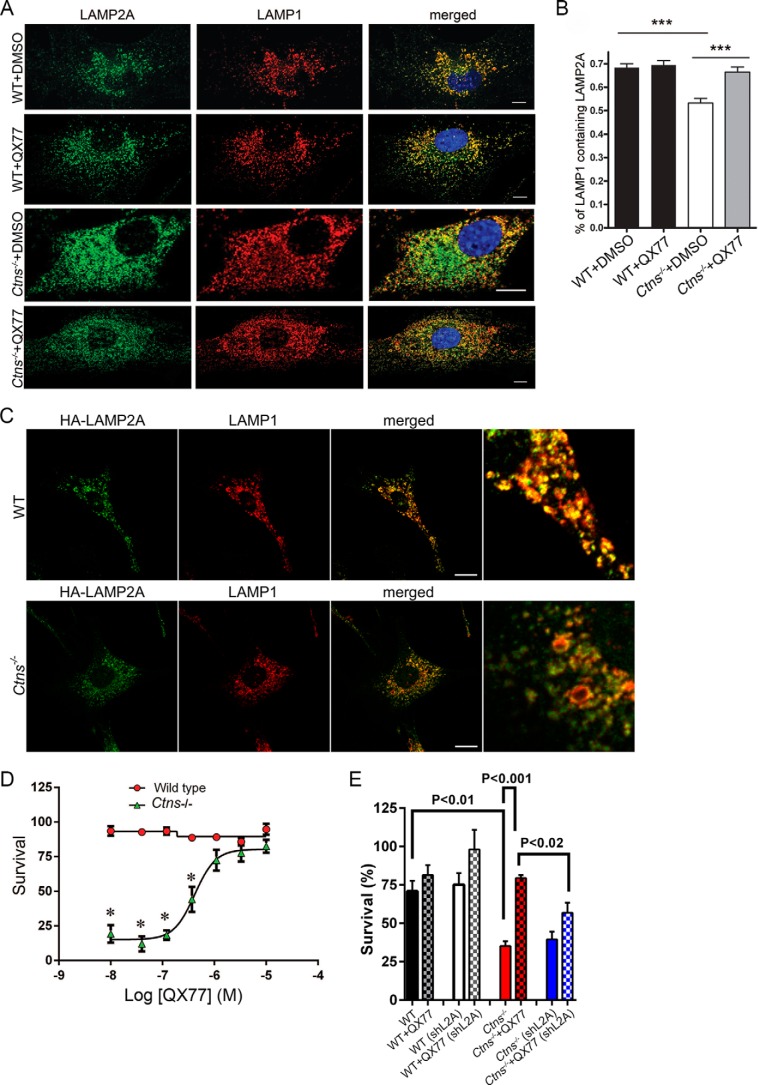
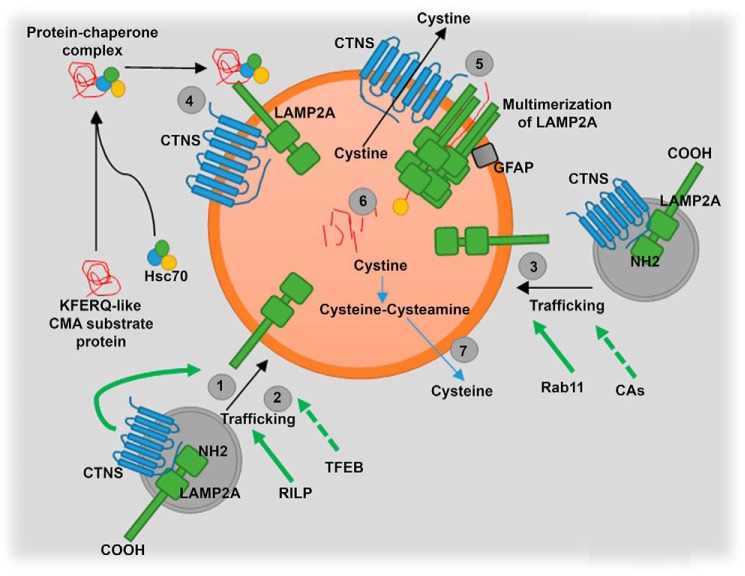
The lysosomal storage disease cystinosis, caused by cystinosin deficiency, is characterized by cell malfunction, tissue failure, and progressive renal injury despite cystine-depletion therapies. Cystinosis is associated with defects in chaperone-mediated autophagy (CMA), but the molecular mechanisms are incompletely understood. Here, we show CMA substrate accumulation in cystinotic kidney proximal tubule cells. We also found mislocalization of the CMA lysosomal receptor LAMP2A and impaired substrate translocation into the lysosome caused by defective CMA in cystinosis. The impaired LAMP2A trafficking and localization were rescued either by the expression of wild-type cystinosin or by the disease-associated point mutant CTNS-K280R, which has no cystine transporter activity. Defective LAMP2A trafficking in cystinosis was found to associate with decreased expression of the small GTPase Rab11 and the Rab7 effector RILP. Defective Rab11 trafficking in cystinosis was rescued by treatment with small-molecule CMA activators. RILP expression was restored by up-regulation of the transcription factor EB (TFEB), which was down-regulated in cystinosis. Although LAMP2A expression is independent of TFEB, TFEB up-regulation corrected lysosome distribution and lysosomal LAMP2A localization in Ctns-/- cells but not Rab11 defects. The up-regulation of Rab11, Rab7, or RILP, but not its truncated form RILP-C33, rescued LAMP2A-defective trafficking in cystinosis, whereas dominant-negative Rab11 or Rab7 impaired LAMP2A trafficking. Treatment of cystinotic cells with a CMA activator increased LAMP2A localization at the lysosome and increased cell survival. Altogether, we show that LAMP2A trafficking is regulated by cystinosin, Rab11, and RILP and that CMA up-regulation is a potential clinically relevant mechanism to increase cell survival in cystinosis.
Keywords: autophagy; cell biology; lysosomal storage disease; lysosome; membrane trafficking.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
-
- Town M., Jean G., Cherqui S., Attard M., Forestier L., Whitmore S. A., Callen D. F., Gribouval O., Broyer M., Bates G. P., van't Hoff W., and Antignac C. (1998) A novel gene encoding an integral membrane protein is mutated in nephropathic cystinosis. Nat. Genet. 18, 319–324 - PubMed
-
- Gahl W. A., Thoene J. G., and Schneider J. A. (2002) Cystinosis. N. Engl. J. Med. 347, 111–121 - PubMed
-
- Kalatzis V., Nevo N., Cherqui S., Gasnier B., and Antignac C. (2004) Molecular pathogenesis of cystinosis: effect of CTNS mutations on the transport activity and subcellular localization of cystinosin. Hum. Mol. Genet. 13, 1361–1371 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
