

Elevated FOXG1 and SOX2 in glioblastoma enforces neural stem cell identity through transcriptional control of cell cycle and epigenetic regulators
- PMID: 28465359
- PMCID: PMC5435889
- DOI: 10.1101/gad.293027.116
Elevated FOXG1 and SOX2 in glioblastoma enforces neural stem cell identity through transcriptional control of cell cycle and epigenetic regulators
Abstract
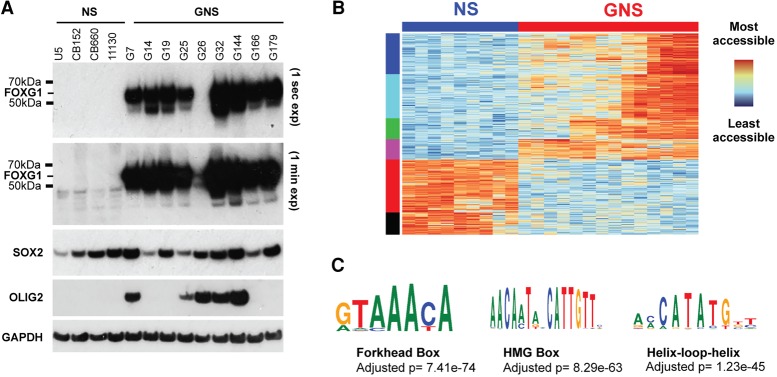
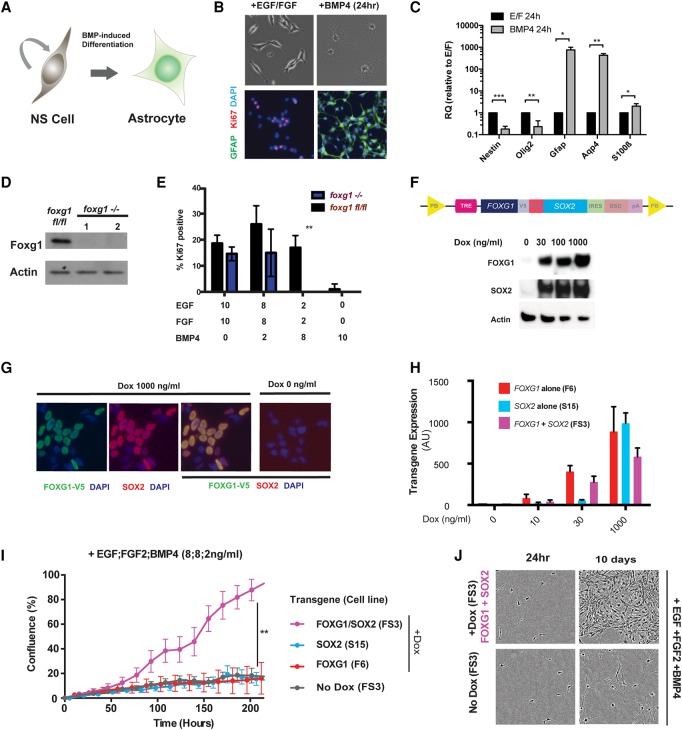
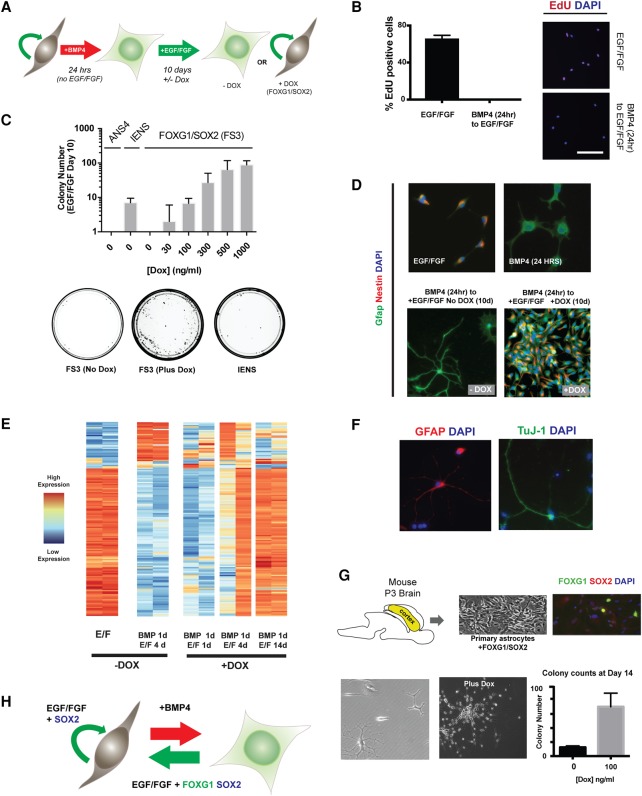
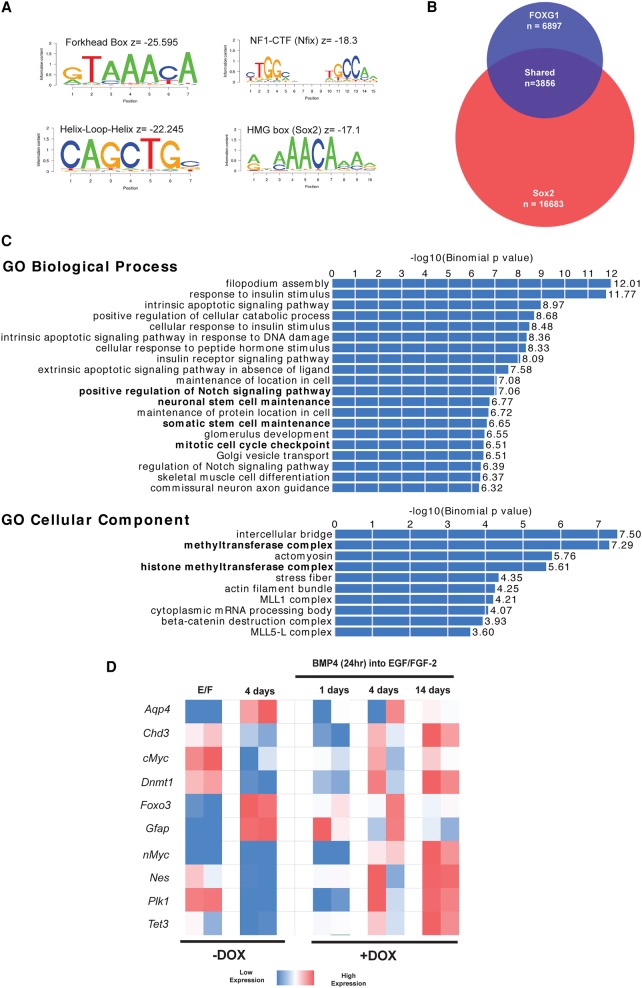
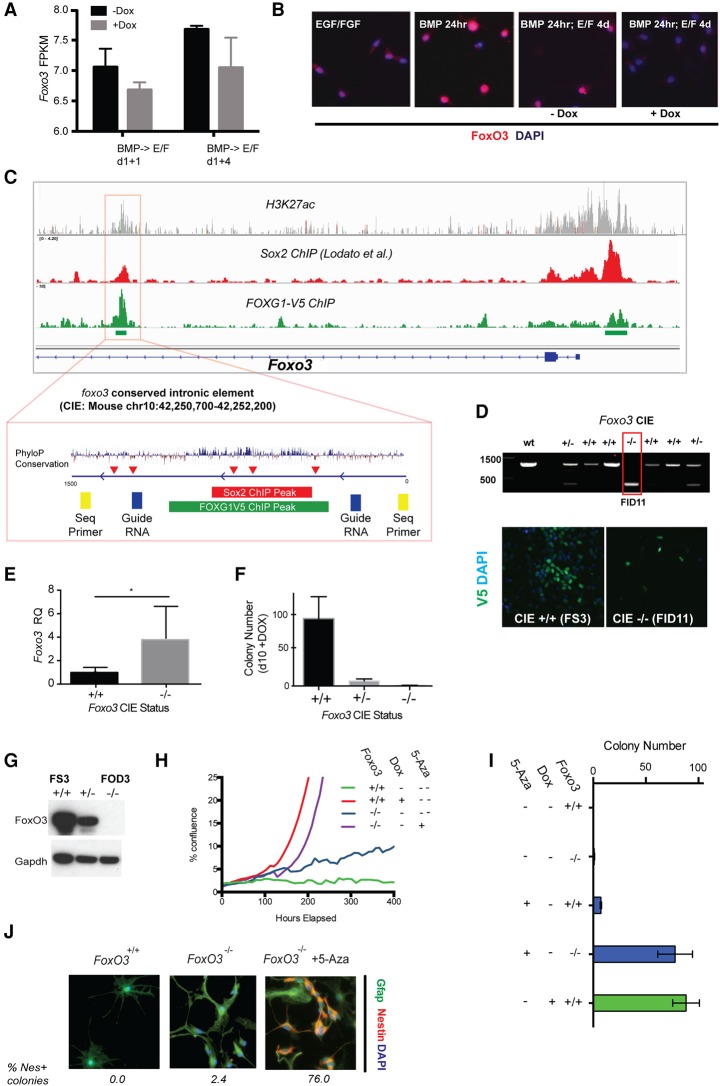
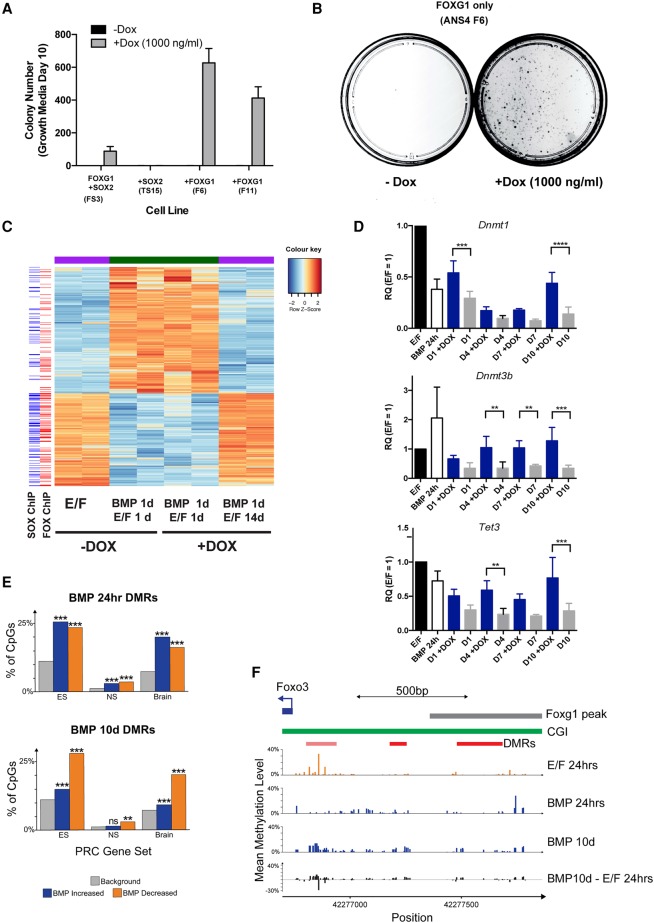
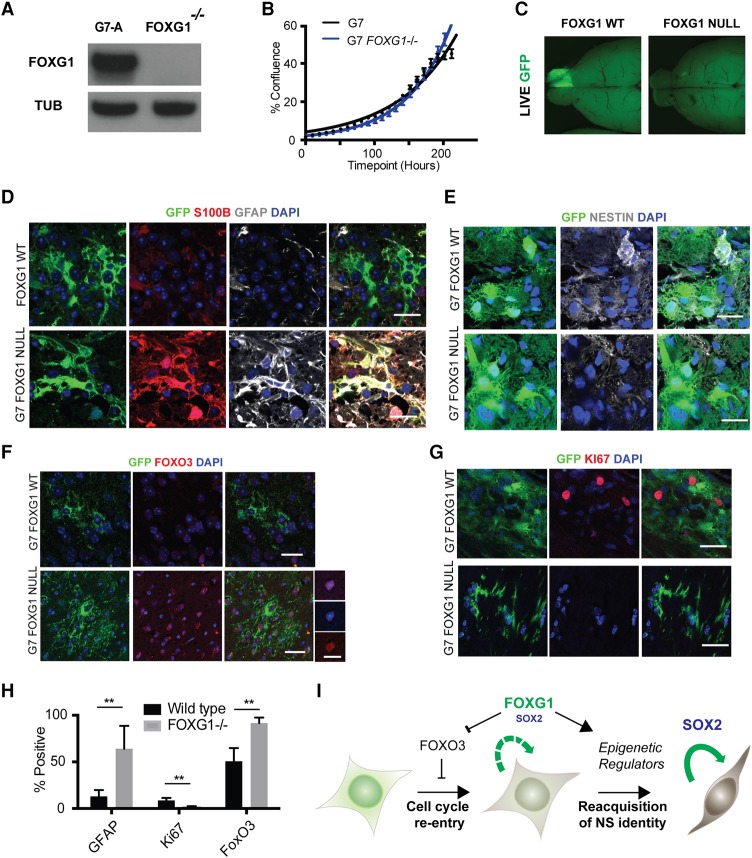
Glioblastoma multiforme (GBM) is an aggressive brain tumor driven by cells with hallmarks of neural stem (NS) cells. GBM stem cells frequently express high levels of the transcription factors FOXG1 and SOX2. Here we show that increased expression of these factors restricts astrocyte differentiation and can trigger dedifferentiation to a proliferative NS cell state. Transcriptional targets include cell cycle and epigenetic regulators (e.g., Foxo3, Plk1, Mycn, Dnmt1, Dnmt3b, and Tet3). Foxo3 is a critical repressed downstream effector that is controlled via a conserved FOXG1/SOX2-bound cis-regulatory element. Foxo3 loss, combined with exposure to the DNA methylation inhibitor 5-azacytidine, enforces astrocyte dedifferentiation. DNA methylation profiling in differentiating astrocytes identifies changes at multiple polycomb targets, including the promoter of Foxo3 In patient-derived GBM stem cells, CRISPR/Cas9 deletion of FOXG1 does not impact proliferation in vitro; however, upon transplantation in vivo, FOXG1-null cells display increased astrocyte differentiation and up-regulate FOXO3. In contrast, SOX2 ablation attenuates proliferation, and mutant cells cannot be expanded in vitro. Thus, FOXG1 and SOX2 operate in complementary but distinct roles to fuel unconstrained self-renewal in GBM stem cells via transcriptional control of core cell cycle and epigenetic regulators.
Keywords: astrocyte; cell cycle; dedifferentiation; epigenetics; glioblastoma; neural stem cell.
© 2017 Bulstrode et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Bachoo RM, Maher EA, Ligon KL, Sharpless NE, Chan SS, You MJ, Tang Y, DeFrances J, Stover E, Weissleder R, et al. 2002. Epidermal growth factor receptor and Ink4a/Arf: convergent mechanisms governing terminal differentiation and transformation along the neural stem cell to astrocyte axis. Cancer Cell 1: 269–277. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 17368/CRUK_/Cancer Research UK/United Kingdom
- MR/J013137/1/MRC_/Medical Research Council/United Kingdom
- 20837/CRUK_/Cancer Research UK/United Kingdom
- 21992/CRUK_/Cancer Research UK/United Kingdom
- 16399/CRUK_/Cancer Research UK/United Kingdom
- MR/K017047/1/MRC_/Medical Research Council/United Kingdom
- WT_/Wellcome Trust/United Kingdom
- A17368/CRUK_/Cancer Research UK/United Kingdom
- C47648/A20837/CRUK_/Cancer Research UK/United Kingdom
- MC_PC_12009/MRC_/Medical Research Council/United Kingdom
- A19680/CRUK_/Cancer Research UK/United Kingdom
- BB/M018040/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous