

A Polymerization-Associated Structural Switch in FtsZ That Enables Treadmilling of Model Filaments
- PMID: 28465423
- PMCID: PMC5414002
- DOI: 10.1128/mBio.00254-17
A Polymerization-Associated Structural Switch in FtsZ That Enables Treadmilling of Model Filaments
Abstract
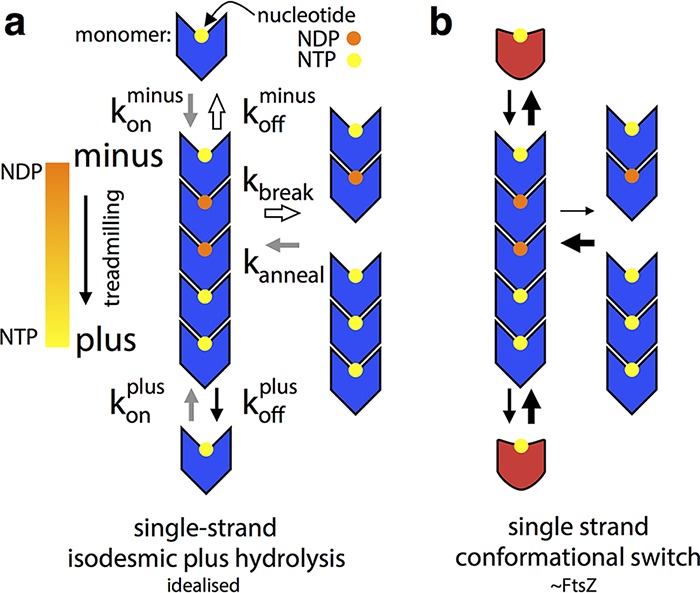
Bacterial cell division in many organisms involves a constricting cytokinetic ring that is orchestrated by the tubulin-like protein FtsZ. FtsZ forms dynamic filaments close to the membrane at the site of division that have recently been shown to treadmill around the division ring, guiding septal wall synthesis. Here, using X-ray crystallography of Staphylococcus aureus FtsZ (SaFtsZ), we reveal how an FtsZ can adopt two functionally distinct conformations, open and closed. The open form is found in SaFtsZ filaments formed in crystals and also in soluble filaments of Escherichia coli FtsZ as deduced by electron cryomicroscopy. The closed form is found within several crystal forms of two nonpolymerizing SaFtsZ mutants and corresponds to many previous FtsZ structures from other organisms. We argue that FtsZ's conformational switch is polymerization-associated, driven by the formation of the longitudinal intersubunit interfaces along the filament. We show that such a switch provides explanations for both how treadmilling may occur within a single-stranded filament and why filament assembly is cooperative.IMPORTANCE The FtsZ protein is a key molecule during bacterial cell division. FtsZ forms filaments that organize cell membrane constriction, as well as remodeling of the cell wall, to divide cells. FtsZ functions through nucleotide-driven filament dynamics that are poorly understood at the molecular level. In particular, mechanisms for cooperative assembly (nonlinear dependency on concentration) and treadmilling (preferential growth at one filament end and loss at the other) have remained elusive. Here, we show that most likely all FtsZ proteins have two distinct conformations, a "closed" form in monomeric FtsZ and an "open" form in filaments. The conformational switch that occurs upon polymerization explains cooperativity and, in concert with polymerization-dependent nucleotide hydrolysis, efficient treadmilling of FtsZ polymers.
Copyright © 2017 Wagstaff et al.
Figures






Similar articles
-
FtsZ filaments have the opposite kinetic polarity of microtubules.Proc Natl Acad Sci U S A. 2018 Oct 16;115(42):10768-10773. doi: 10.1073/pnas.1811919115. Epub 2018 Oct 1. Proc Natl Acad Sci U S A. 2018. PMID: 30275322 Free PMC article.
-
Nucleotide-induced folding of cell division protein FtsZ from Staphylococcus aureus.FEBS J. 2020 Sep;287(18):4048-4067. doi: 10.1111/febs.15235. Epub 2020 Feb 24. FEBS J. 2020. PMID: 31997533
-
Identification of the key interactions in structural transition pathway of FtsZ from Staphylococcus aureus.J Struct Biol. 2017 May;198(2):65-73. doi: 10.1016/j.jsb.2017.04.008. Epub 2017 Apr 27. J Struct Biol. 2017. PMID: 28456664
-
At the Heart of Bacterial Cytokinesis: The Z Ring.Trends Microbiol. 2019 Sep;27(9):781-791. doi: 10.1016/j.tim.2019.04.011. Epub 2019 Jun 3. Trends Microbiol. 2019. PMID: 31171437 Free PMC article. Review.
-
Insights into the Structure, Function, and Dynamics of the Bacterial Cytokinetic FtsZ-Ring.Annu Rev Biophys. 2020 May 6;49:309-341. doi: 10.1146/annurev-biophys-121219-081703. Epub 2020 Feb 24. Annu Rev Biophys. 2020. PMID: 32092282 Free PMC article. Review.
Cited by
-
An essential Staphylococcus aureus cell division protein directly regulates FtsZ dynamics.Elife. 2018 Oct 2;7:e38856. doi: 10.7554/eLife.38856. Elife. 2018. PMID: 30277210 Free PMC article.
-
High-resolution crystal structures of Escherichia coli FtsZ bound to GDP and GTP.Acta Crystallogr F Struct Biol Commun. 2020 Feb 1;76(Pt 2):94-102. doi: 10.1107/S2053230X20001132. Epub 2020 Feb 5. Acta Crystallogr F Struct Biol Commun. 2020. PMID: 32039891 Free PMC article.
-
MinC and FtsZ mutant analysis provides insight into MinC/MinD-mediated Z ring disassembly.J Biol Chem. 2018 Apr 20;293(16):5834-5846. doi: 10.1074/jbc.M117.815894. Epub 2018 Feb 2. J Biol Chem. 2018. PMID: 29414773 Free PMC article.
-
FtsZ phosphorylation brings about growth arrest upon DNA damage in Deinococcus radiodurans.FASEB Bioadv. 2022 Oct 31;5(1):27-42. doi: 10.1096/fba.2022-00082. eCollection 2023 Jan. FASEB Bioadv. 2022. PMID: 36643897 Free PMC article.
-
Crystal structures of the cell-division protein FtsZ from Klebsiella pneumoniae and Escherichia coli.Acta Crystallogr F Struct Biol Commun. 2020 Feb 1;76(Pt 2):86-93. doi: 10.1107/S2053230X2000076X. Epub 2020 Feb 5. Acta Crystallogr F Struct Biol Commun. 2020. PMID: 32039890 Free PMC article.
