

Redesigning the QA binding site of Photosystem II allows reduction of exogenous quinones
- PMID: 28466860
- PMCID: PMC5418674
- DOI: 10.1038/ncomms15274
Redesigning the QA binding site of Photosystem II allows reduction of exogenous quinones
Abstract
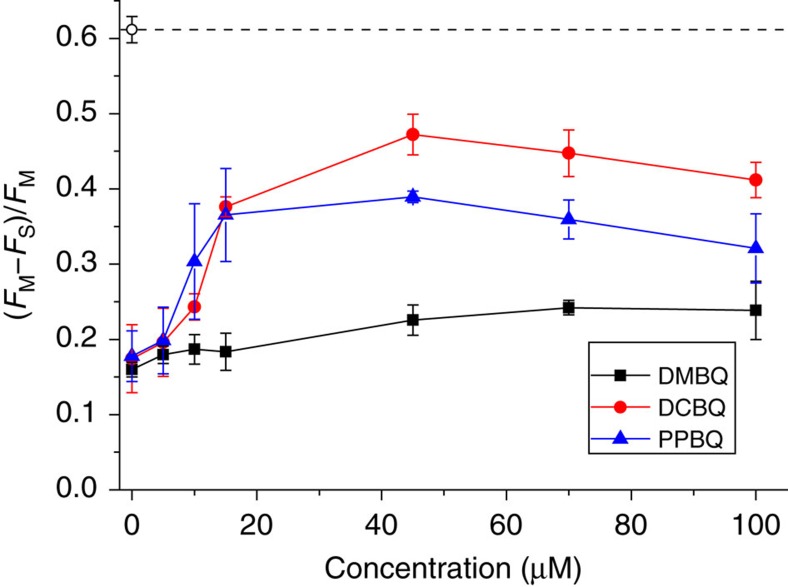
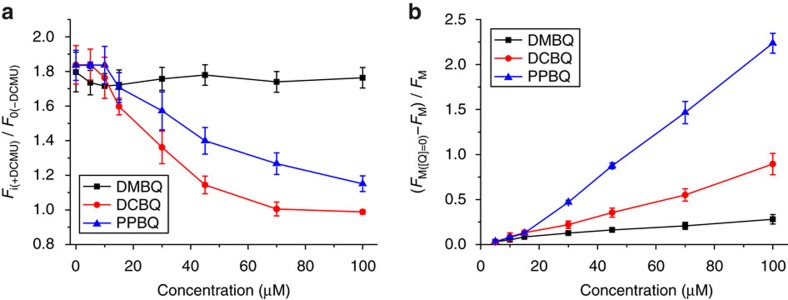
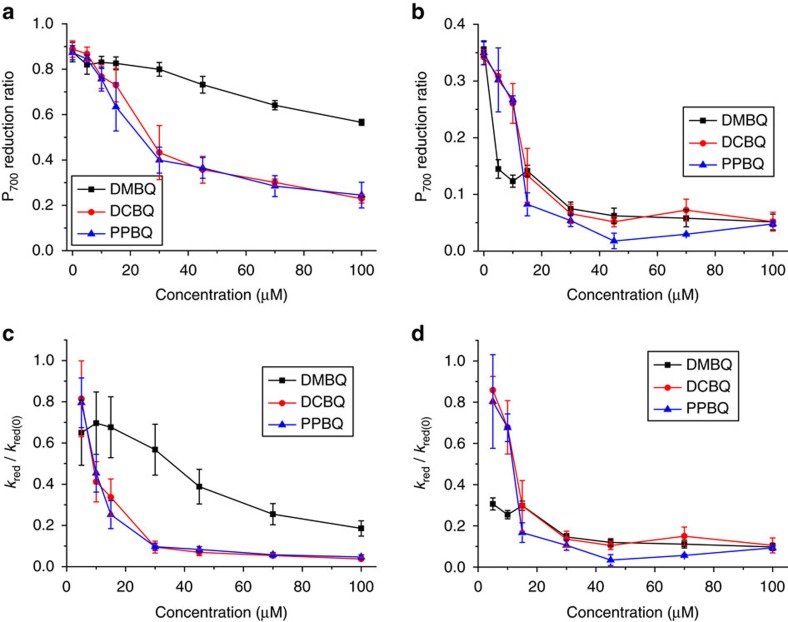
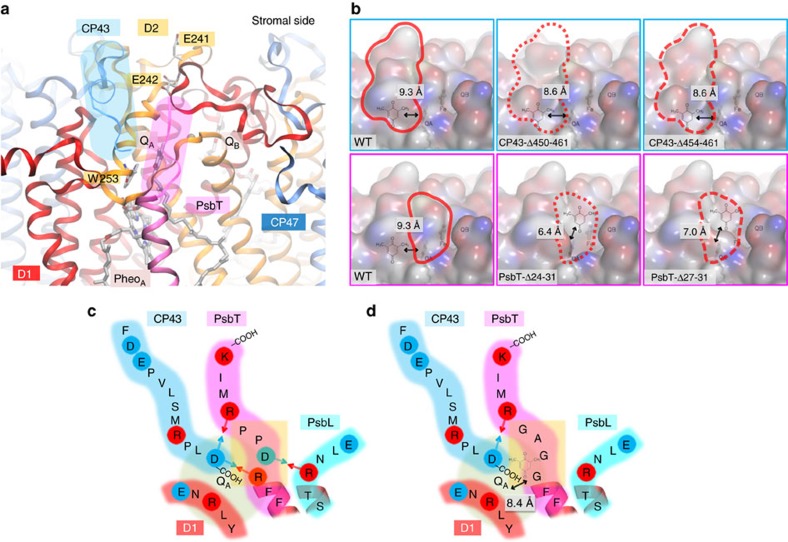
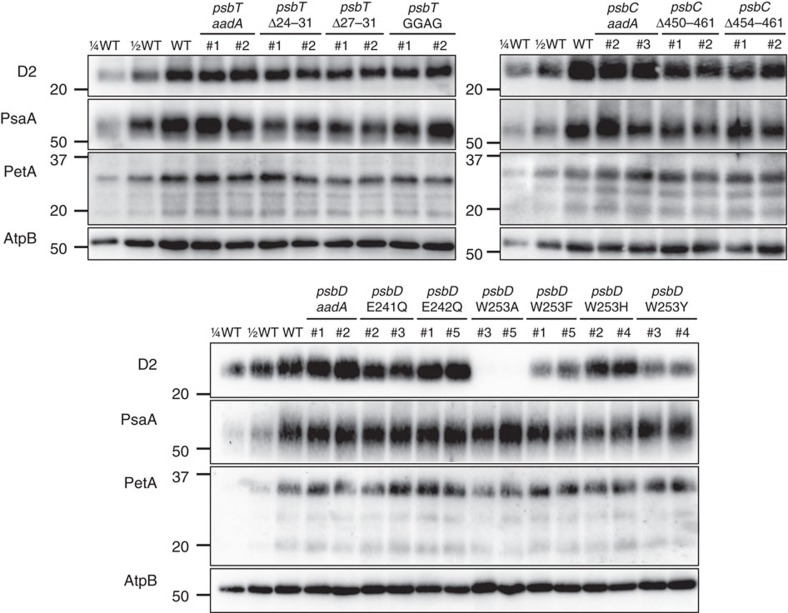
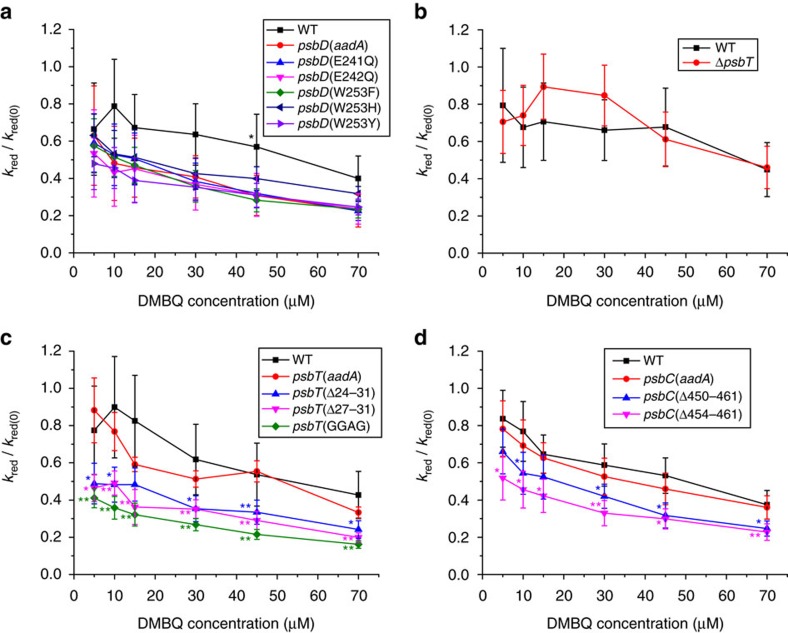
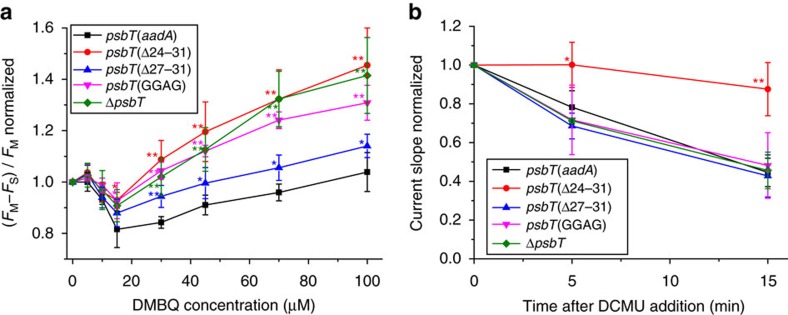
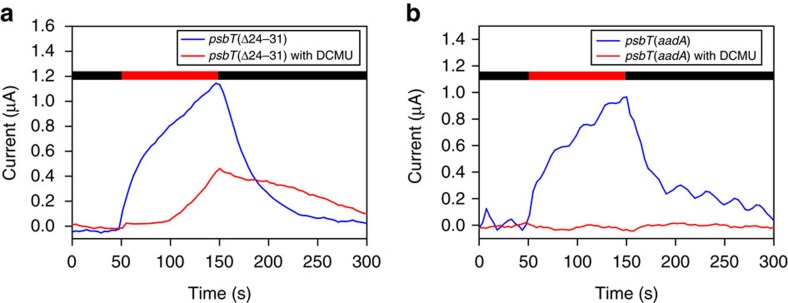
Strategies to harness photosynthesis from living organisms to generate electrical power have long been considered, yet efficiency remains low. Here, we aimed to reroute photosynthetic electron flow in photosynthetic organisms without compromising their phototrophic properties. We show that 2,6-dimethyl-p-benzoquinone (DMBQ) can be used as an electron mediator to assess the efficiency of mutations designed to engineer a novel electron donation pathway downstream of the primary electron acceptor QA of Photosystem (PS) II in the green alga Chlamydomonas reinhardtii. Through the use of structural prediction studies and a screen of site-directed PSII mutants we show that modifying the environment of the QA site increases the reduction rate of DMBQ. Truncating the C-terminus of the PsbT subunit protruding in the stroma provides evidence that shortening the distance between QA and DMBQ leads to sustained electron transfer to DMBQ, as confirmed by chronoamperometry, consistent with a bypass of the natural QA°- to QB pathway.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Rosenbaum M., He Z. & Angenent L. T. Light energy to bioelectricity: photosynthetic microbial fuel cells. Curr. Opin. Biotechnol. 21, 259–264 (2010). - PubMed
-
- McCormick A. J. et al.. Biophotovoltaics: oxygenic photosynthetic organisms in the world of bioelectrochemical systems. Energy Environ. Sci. 8, 1092–1109 (2015).
-
- Sekar N. & Ramasamy R. P. Recent advances in photosynthetic energy conversion. J. Photochem. Photobiol. C: Photochem. Rev. 22, 19–33 (2015).
-
- Wraight C. A. & Clayton R. K. The absolute quantum efficiency of bacteriochlorophyll photooxidation in reaction centres of Rhodopseudomonas spheroides. Biochim. Biophys. Acta 333, 246–260 (1974). - PubMed
-
- Björkman O. & Demmig B. Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta 170, 489–504 (1987). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
