

Redox Signaling in Neurotransmission and Cognition During Aging
- PMID: 28467718
- PMCID: PMC5962336
- DOI: 10.1089/ars.2017.7111
Redox Signaling in Neurotransmission and Cognition During Aging
Abstract
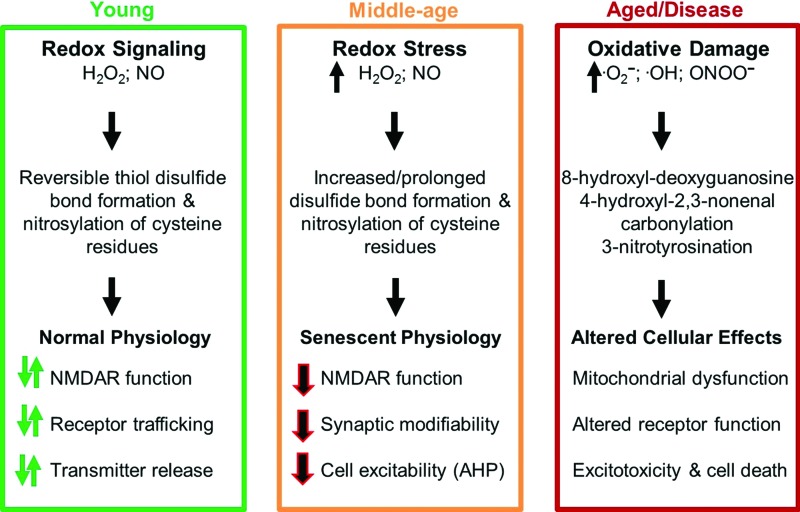
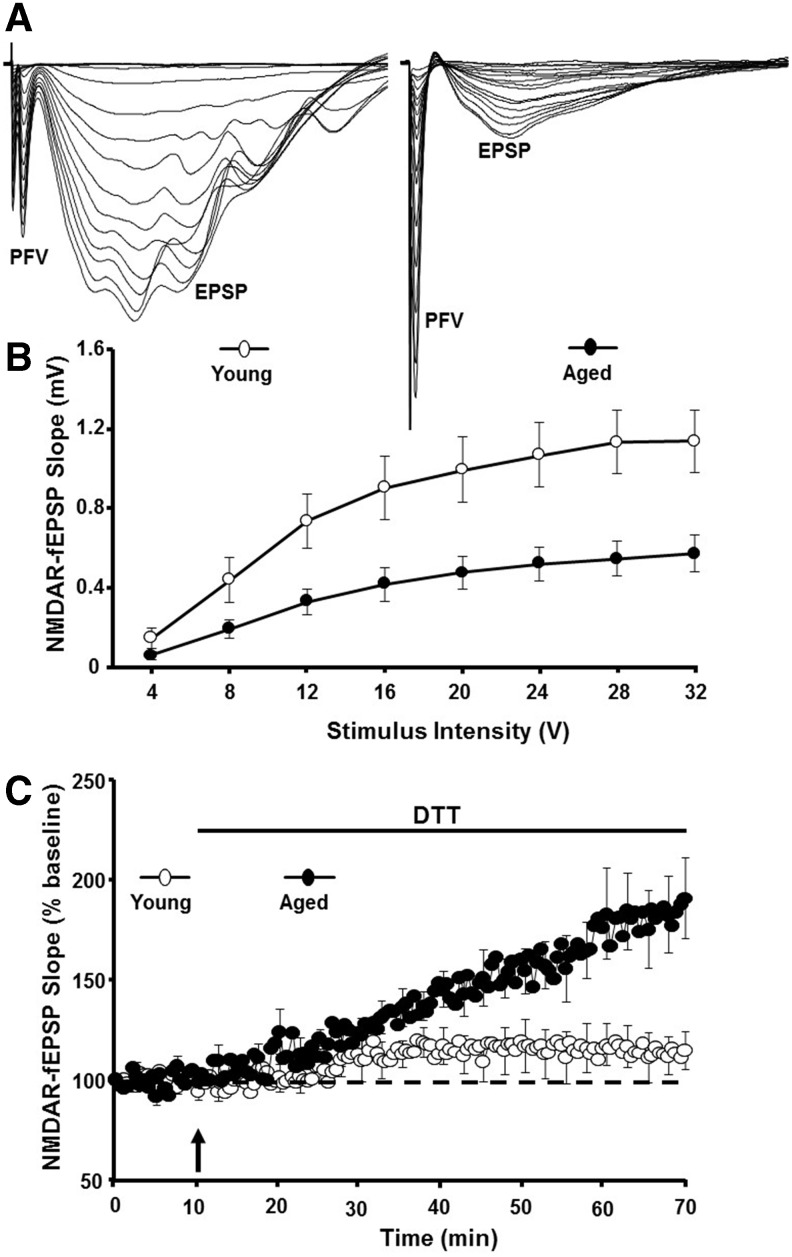
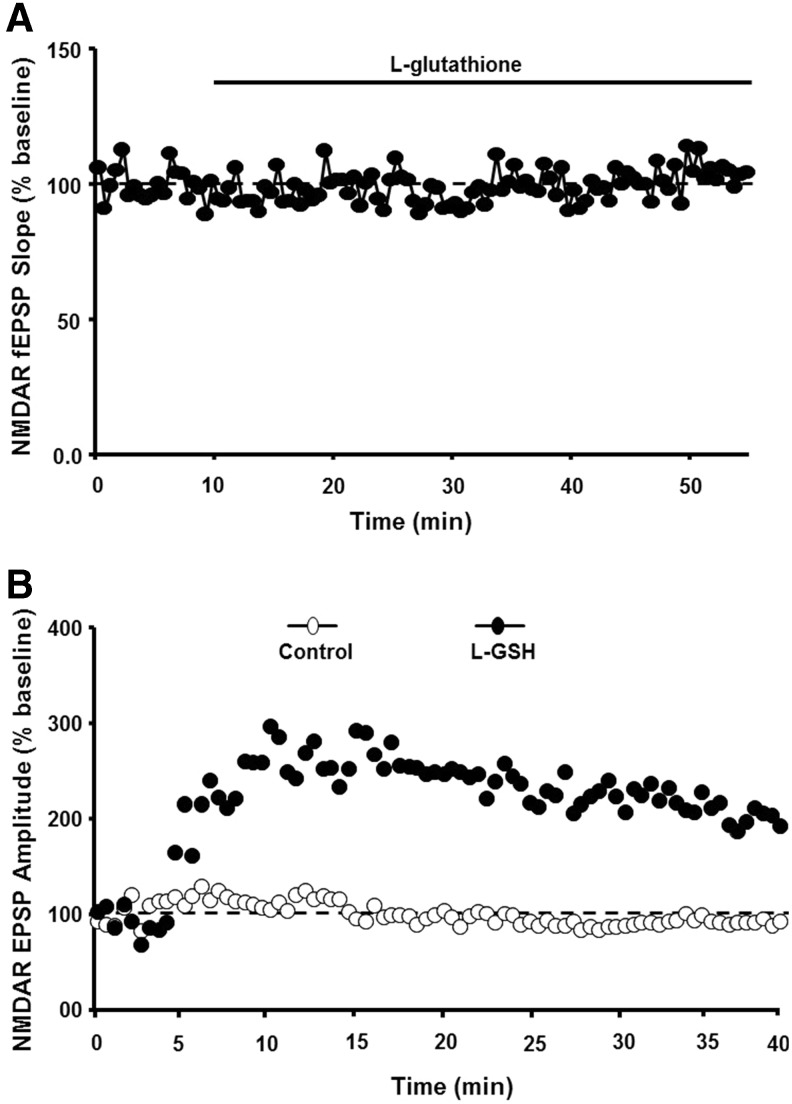
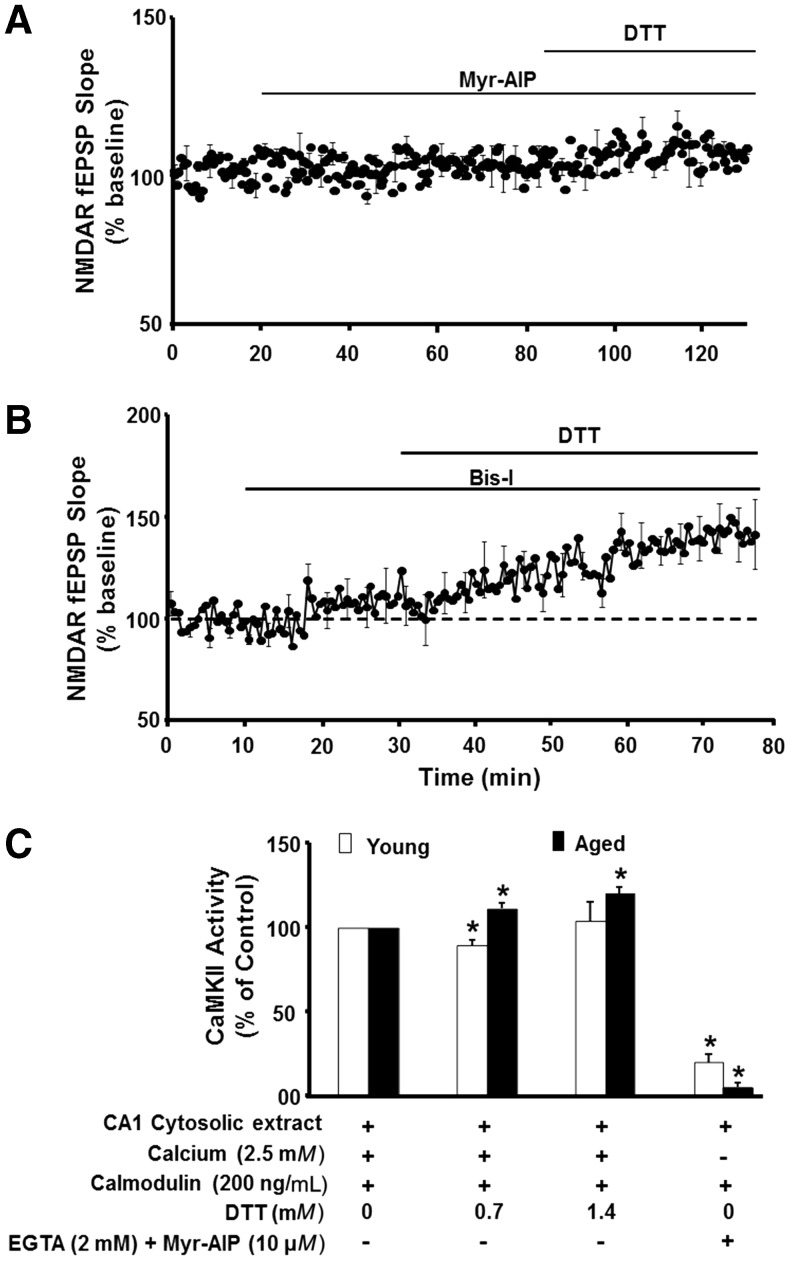
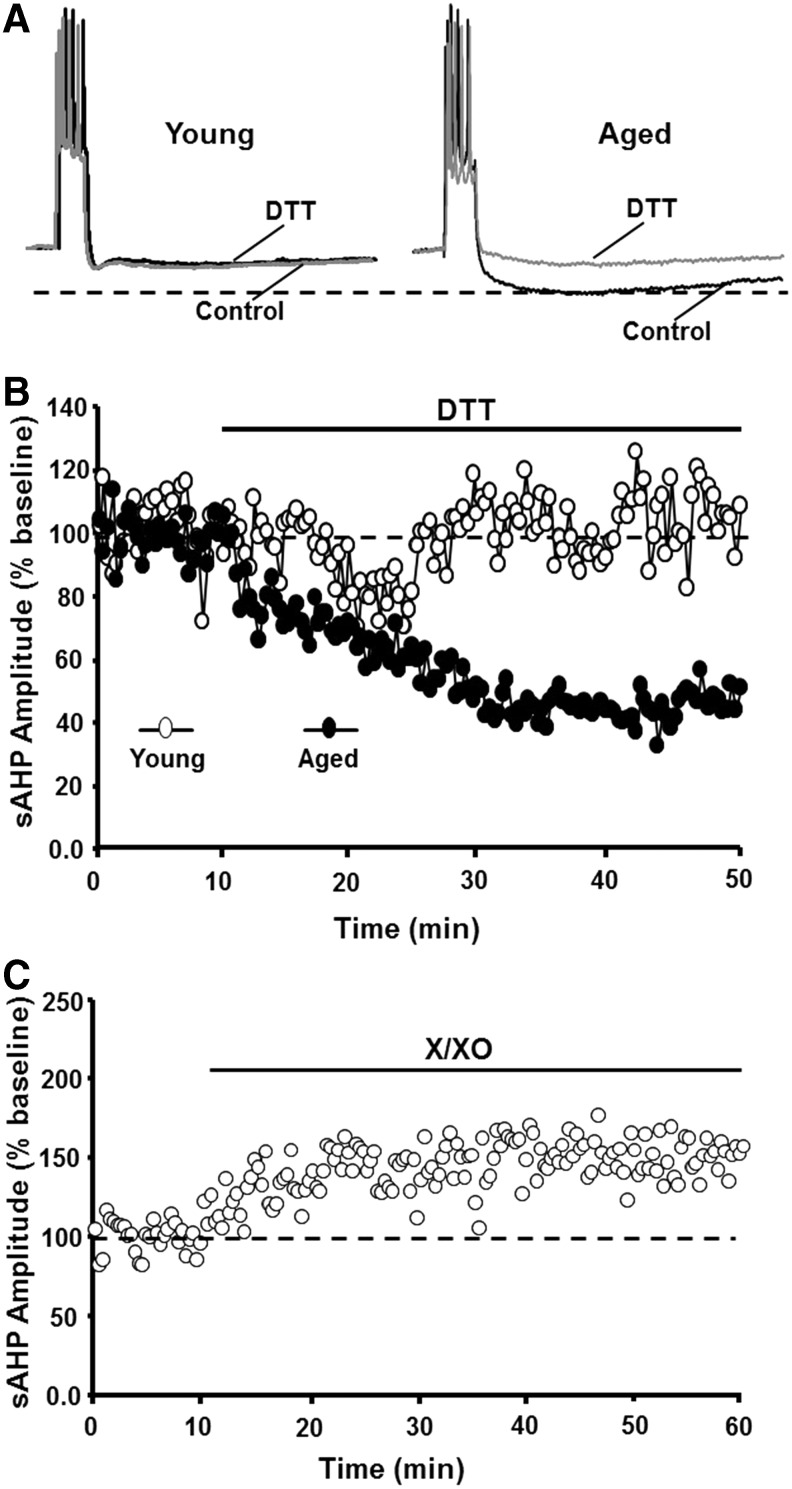
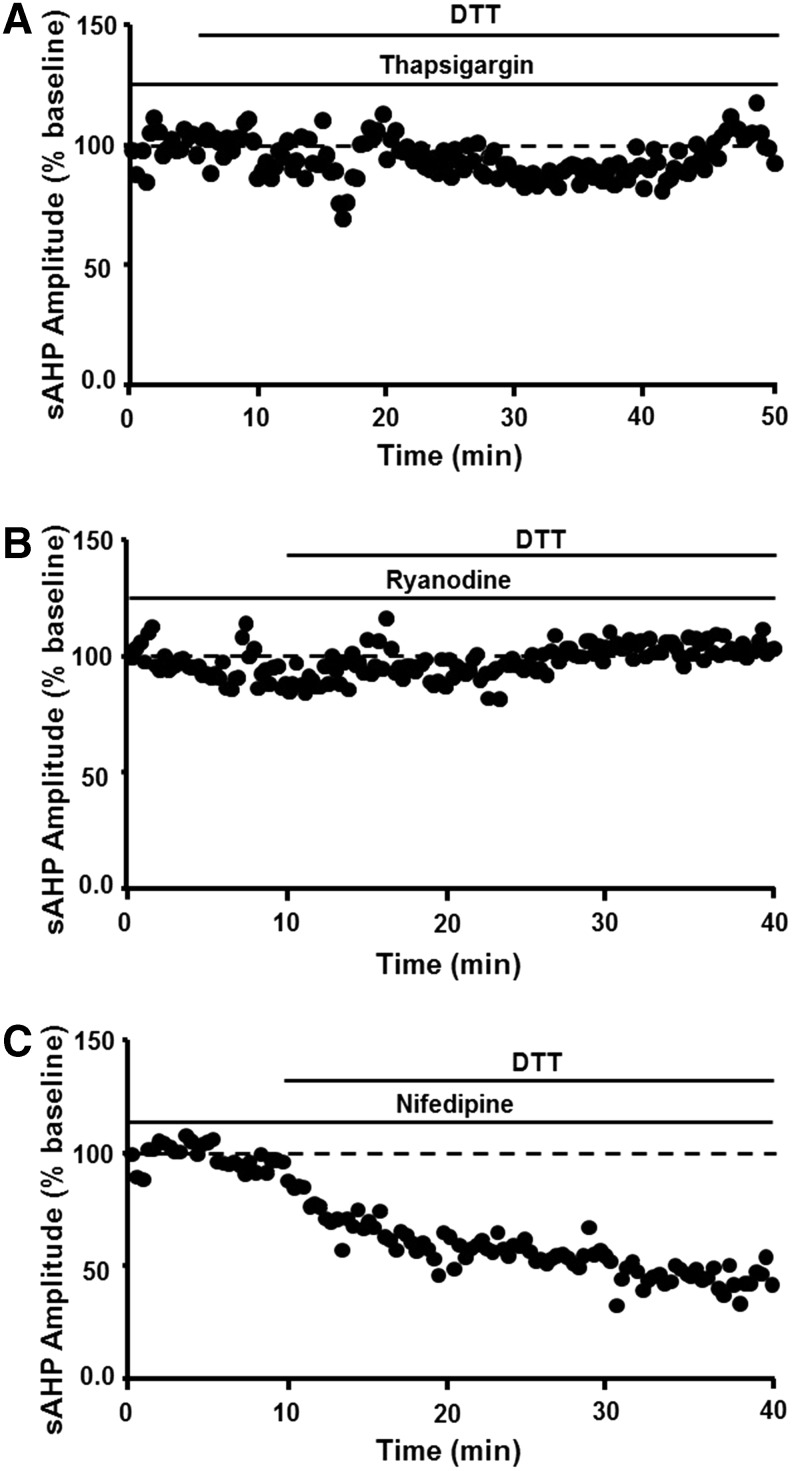
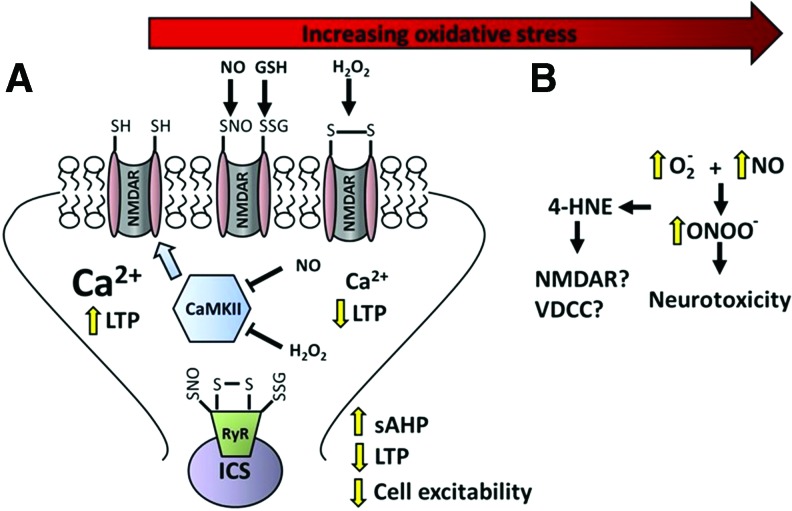
Significance: Oxidative stress increases in the brain with aging and neurodegenerative diseases. Previous work emphasized irreversible oxidative damage in relation to cognitive impairment. This research has evolved to consider a continuum of alterations, from redox signaling to oxidative damage, which provides a basis for understanding the onset and progression of cognitive impairment. This review provides an update on research linking redox signaling to altered function of neural circuits involved in information processing and memory. Recent Advances: Starting in middle age, redox signaling triggers changes in nervous system physiology described as senescent physiology. Hippocampal senescent physiology involves decreased cell excitability, altered synaptic plasticity, and decreased synaptic transmission. Recent studies indicate N-methyl-d-aspartate and ryanodine receptors and Ca2+ signaling molecules as molecular substrates of redox-mediated senescent physiology.
Critical issues: We review redox homeostasis mechanisms and consider the chemical character of reactive oxygen and nitrogen species and their role in regulating different transmitter systems. In this regard, senescent physiology may represent the co-opting of pathways normally responsible for feedback regulation of synaptic transmission. Furthermore, differences across transmitter systems may underlie differential vulnerability of brain regions and neuronal circuits to aging and disease.
Future directions: It will be important to identify the intrinsic mechanisms for the shift in oxidative/reductive processes. Intrinsic mechanism will depend on the transmitter system, oxidative stressors, and expression/activity of antioxidant enzymes. In addition, it will be important to identify how intrinsic processes interact with other aging factors, including changes in inflammatory or hormonal signals. Antioxid. Redox Signal. 28, 1724-1745.
Keywords: Ca2+ signaling; NMDA receptor; aging; cognition; oxidative stress; redox regulation.
Figures

Similar articles
-
Central role for NMDA receptors in redox mediated impairment of synaptic function during aging and Alzheimer's disease.Behav Brain Res. 2017 Mar 30;322(Pt B):223-232. doi: 10.1016/j.bbr.2016.05.012. Epub 2016 May 11. Behav Brain Res. 2017. PMID: 27180169 Review.
-
Redox Biology in Neurological Function, Dysfunction, and Aging.Antioxid Redox Signal. 2018 Jun 20;28(18):1583-1586. doi: 10.1089/ars.2018.7509. Epub 2018 Apr 23. Antioxid Redox Signal. 2018. PMID: 29634346 Free PMC article.
-
Role of antioxidant enzymes in redox regulation of N-methyl-D-aspartate receptor function and memory in middle-aged rats.Neurobiol Aging. 2014 Jun;35(6):1459-68. doi: 10.1016/j.neurobiolaging.2013.12.002. Epub 2013 Dec 9. Neurobiol Aging. 2014. PMID: 24388786 Free PMC article.
-
Oxidative stress and protein aggregation during biological aging.Exp Gerontol. 2001 Sep;36(9):1539-50. doi: 10.1016/s0531-5565(01)00139-5. Exp Gerontol. 2001. PMID: 11525876 Review.
-
Contribution of Ca2+ release channels to hippocampal synaptic plasticity and spatial memory: potential redox modulation.Antioxid Redox Signal. 2014 Aug 20;21(6):892-914. doi: 10.1089/ars.2013.5796. Epub 2014 Mar 11. Antioxid Redox Signal. 2014. PMID: 24410659 Review.
Cited by
-
ERRα regulates synaptic transmission through reactive oxygen species in hippocampal neurons.Heliyon. 2023 Dec 15;10(1):e23739. doi: 10.1016/j.heliyon.2023.e23739. eCollection 2024 Jan 15. Heliyon. 2023. PMID: 38192817 Free PMC article.
-
Subunit contribution to NMDA receptor hypofunction and redox sensitivity of hippocampal synaptic transmission during aging.Aging (Albany NY). 2019 Jul 24;11(14):5140-5157. doi: 10.18632/aging.102108. Aging (Albany NY). 2019. PMID: 31339863 Free PMC article.
-
Sex Differences in Metabolic Indices and Chronic Neuroinflammation in Response to Prolonged High-Fat Diet in ApoE4 Knock-In Mice.Int J Mol Sci. 2022 Apr 1;23(7):3921. doi: 10.3390/ijms23073921. Int J Mol Sci. 2022. PMID: 35409283 Free PMC article.
-
The Puzzling Role of Neuron-Specific PMCA Isoforms in the Aging Process.Int J Mol Sci. 2019 Dec 16;20(24):6338. doi: 10.3390/ijms20246338. Int J Mol Sci. 2019. PMID: 31888192 Free PMC article. Review.
-
Oxidative Stress-Related Mechanisms in Schizophrenia Pathogenesis and New Treatment Perspectives.Oxid Med Cell Longev. 2021 Jan 23;2021:8881770. doi: 10.1155/2021/8881770. eCollection 2021. Oxid Med Cell Longev. 2021. PMID: 33552387 Free PMC article. Review.
References
-
- Abd El Mohsen MM, Iravani MM, Spencer JP, Rose S, Fahim AT, Motawi TM, Ismail NA, and Jenner P. Age-associated changes in protein oxidation and proteasome activities in rat brain: Modulation by antioxidants. Biochem Biophys Res Commun 336: 386–391, 2005 - PubMed
-
- Adamczyk A, Kazmierczak A, and Strosznajder JB. Alpha-synuclein and its neurotoxic fragment inhibit dopamine uptake into rat striatal synaptosomes. Relationship to nitric oxide. Neurochem Int 49: 407–412, 2006 - PubMed
-
- Aizenman E, Hartnett KA, and Reynolds IJ. Oxygen free radicals regulate NMDA receptor function via a redox modulatory site. Neuron 5: 841–846, 1990 - PubMed
-
- Aizenman E, Lipton SA, and Loring RH. Selective modulation of NMDA responses by reduction and oxidation. Neuron 2: 1257–1263, 1989 - PubMed
-
- Akaishi T, Nakazawa K, Sato K, Ohno Y, and Ito Y. 4-Hydroxynonenal modulates the long-term potentiation induced by L-type Ca2+ channel activation in the rat dentate gyrus in vitro. Neurosci Lett 370: 155–159, 2004 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous