

The DEAD-box protein DDX43 (HAGE) is a dual RNA-DNA helicase and has a K-homology domain required for full nucleic acid unwinding activity
- PMID: 28468824
- PMCID: PMC5481555
- DOI: 10.1074/jbc.M117.774950
The DEAD-box protein DDX43 (HAGE) is a dual RNA-DNA helicase and has a K-homology domain required for full nucleic acid unwinding activity
Abstract
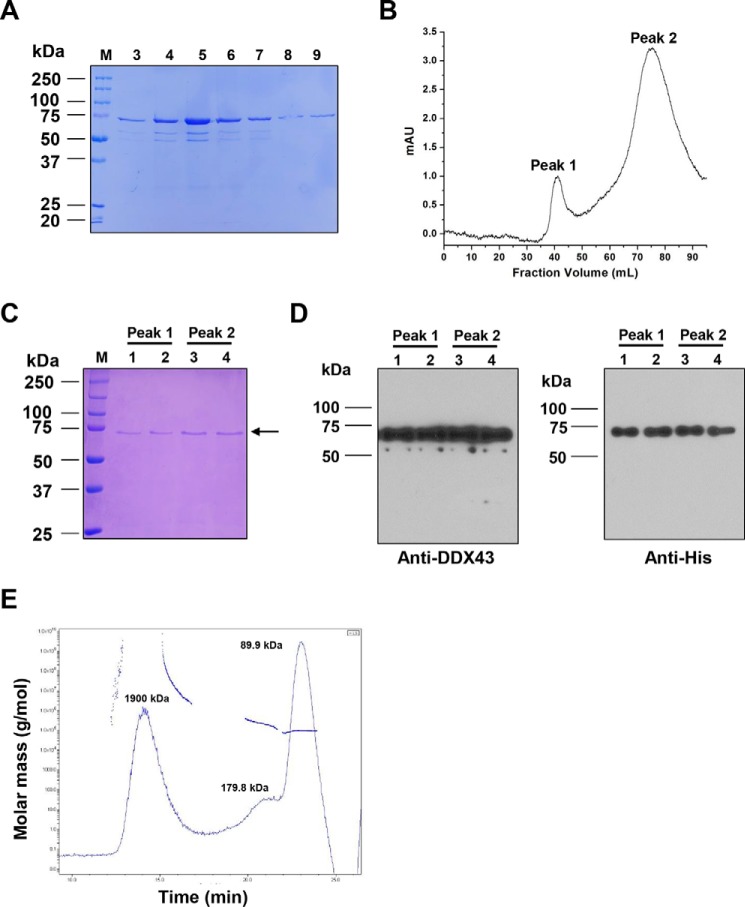
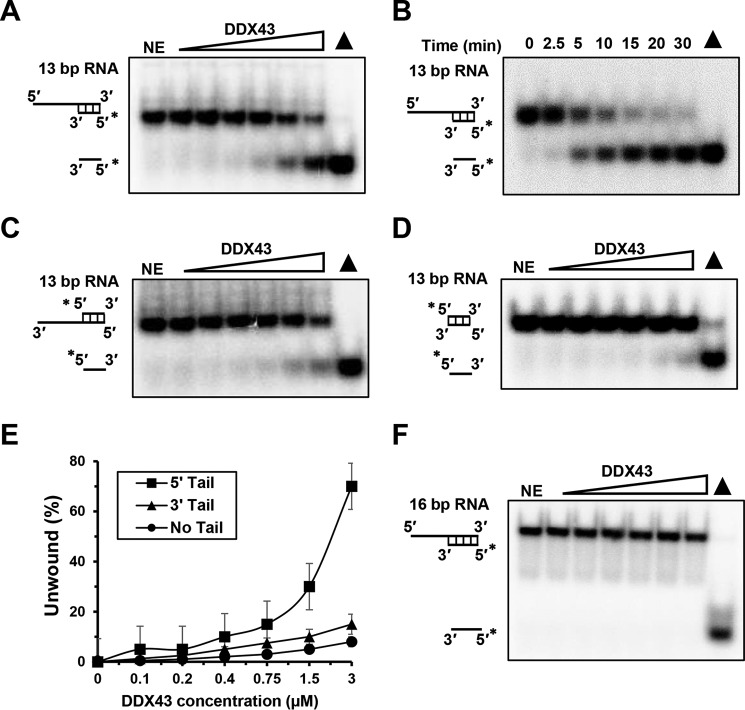
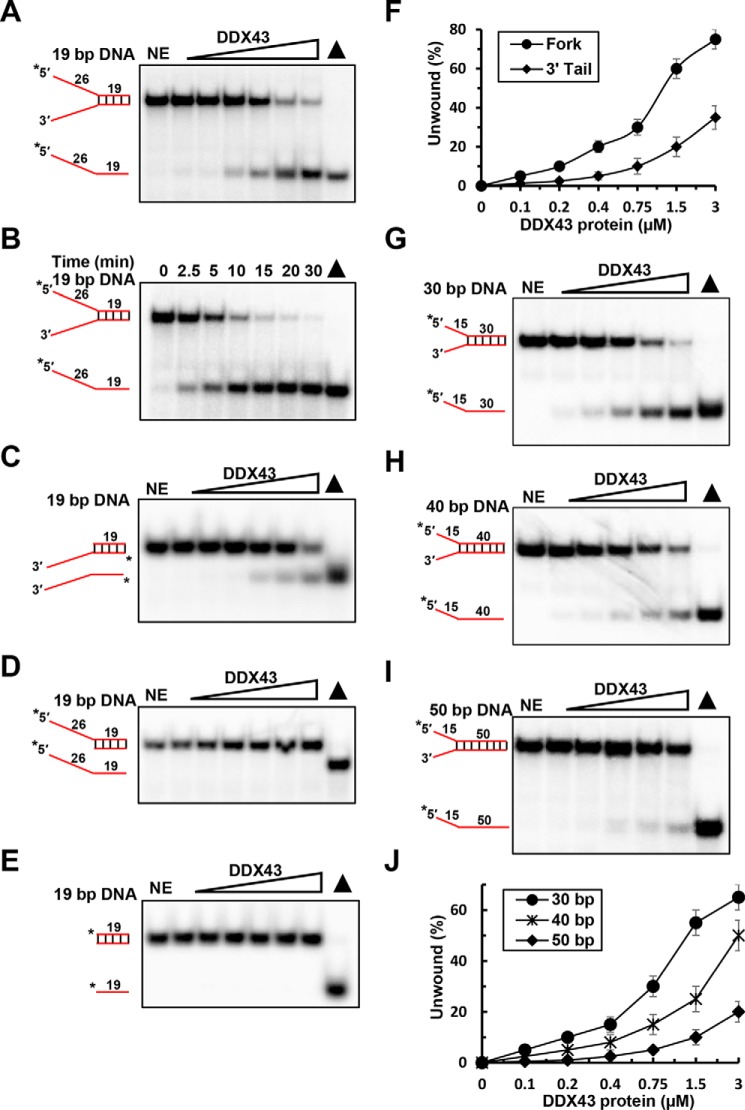
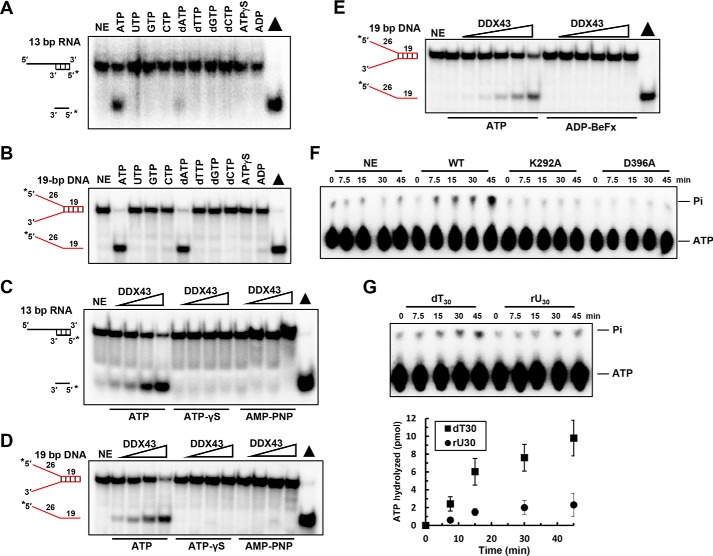
The K-homology (KH) domain is a nucleic acid-binding domain present in many proteins but has not been reported in helicases. DDX43, also known as HAGE (helicase antigen gene), is a member of the DEAD-box protein family. It contains a helicase core domain in its C terminus and a potential KH domain in its N terminus. DDX43 is highly expressed in many tumors and is, therefore, considered a potential target for immunotherapy. Despite its potential as a therapeutic target, little is known about its activities. Here, we purified recombinant DDX43 protein to near homogeneity and found that it exists as a monomer in solution. Biochemical assays demonstrated that it is an ATP-dependent RNA and DNA helicase. Although DDX43 was active on duplex RNA regardless of the orientation of the single-stranded RNA tail, it preferred a 5' to 3' polarity on RNA and a 3' to 5' direction on DNA. Truncation mutations and site-directed mutagenesis confirmed that the KH domain in DDX43 is responsible for nucleic acid binding. Compared with the activity of the full-length protein, the C-terminal helicase domain had no unwinding activity on RNA substrates and had significantly reduced unwinding activity on DNA. Moreover, the full-length DDX43 protein, with single amino acid change in the KH domain, had reduced unwinding and binding activates on RNA and DNA substrates. Our results demonstrate that DDX43 is a dual helicase and the KH domain is required for its full unwinding activity.
Keywords: DNA; DNA enzyme; DNA helicase; DNA-protein interaction; RNA; RNA helicase; RNA metabolism; RNA-binding protein.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
Similar articles
-
The KH domain facilitates the substrate specificity and unwinding processivity of DDX43 helicase.J Biol Chem. 2021 Jan-Jun;296:100085. doi: 10.1074/jbc.RA120.015824. Epub 2020 Nov 23. J Biol Chem. 2021. PMID: 33199368 Free PMC article.
-
DDX43 prefers single strand substrate and its full binding activity requires physical connection of all domains.Biochem Biophys Res Commun. 2019 Dec 10;520(3):594-599. doi: 10.1016/j.bbrc.2019.09.114. Epub 2019 Oct 14. Biochem Biophys Res Commun. 2019. PMID: 31623828
-
Structure-function analysis of DEAD-box helicase DDX43.Methods. 2022 Aug;204:286-299. doi: 10.1016/j.ymeth.2022.03.002. Epub 2022 Mar 4. Methods. 2022. PMID: 35257897
-
The Current View on the Helicase Activity of RNA Helicase A and Its Role in Gene Expression.Curr Protein Pept Sci. 2021;22(1):29-40. doi: 10.2174/1389203721666201103084122. Curr Protein Pept Sci. 2021. PMID: 33143622 Review.
-
The mechanism of ATP-dependent RNA unwinding by DEAD box proteins.Biol Chem. 2009 Dec;390(12):1237-50. doi: 10.1515/BC.2009.135. Biol Chem. 2009. PMID: 19747077 Review.
Cited by
-
DDX RNA helicases: key players in cellular homeostasis and innate antiviral immunity.J Virol. 2024 Oct 22;98(10):e0004024. doi: 10.1128/jvi.00040-24. Epub 2024 Aug 30. J Virol. 2024. PMID: 39212449 Free PMC article. Review.
-
Micronutrient supplementation affects DNA methylation in male gonads with potential intergenerational epigenetic inheritance involving the embryonic development through glutamate receptor-associated genes.BMC Genomics. 2022 Feb 10;23(1):115. doi: 10.1186/s12864-022-08348-4. BMC Genomics. 2022. PMID: 35144563 Free PMC article.
-
Substrate Specificities of DDX1: A Human DEAD-box protein.bioRxiv [Preprint]. 2024 Jan 10:2024.01.09.573566. doi: 10.1101/2024.01.09.573566. bioRxiv. 2024. Update in: ACS Omega. 2025 Jan 14;10(3):2598-2607. doi: 10.1021/acsomega.4c07522. PMID: 38260591 Free PMC article. Updated. Preprint.
-
Consequences and Resolution of Transcription-Replication Conflicts.Life (Basel). 2021 Jun 30;11(7):637. doi: 10.3390/life11070637. Life (Basel). 2021. PMID: 34209204 Free PMC article. Review.
-
The DDX5/Dbp2 subfamily of DEAD-box RNA helicases.Wiley Interdiscip Rev RNA. 2019 Mar;10(2):e1519. doi: 10.1002/wrna.1519. Epub 2018 Dec 2. Wiley Interdiscip Rev RNA. 2019. PMID: 30506978 Free PMC article. Review.
References
-
- Linder P., and Jankowsky E. (2011) From unwinding to clamping: the DEAD box RNA helicase family. Nat. Rev. Mol. Cell Biol. 12, 505–516 - PubMed
-
- Lohman T. M., and Bjornson K. P. (1996) Mechanisms of helicase-catalyzed DNA unwinding. Annu. Rev. Biochem. 65, 169–214 - PubMed
-
- Pyle A. M. (2008) Translocation and unwinding mechanisms of RNA and DNA helicases. Annu. Rev. Biophys. 37, 317–336 - PubMed
-
- Singleton M. R., Dillingham M. S., and Wigley D. B. (2007) Structure and mechanism of helicases and nucleic acid translocases. Annu. Rev. Biochem. 76, 23–50 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
