

GSK-3β Homolog Rim11 and the Histone Deacetylase Complex Ume6-Sin3-Rpd3 Are Involved in Replication Stress Response Caused by Defects in Dna2
- PMID: 28468907
- PMCID: PMC5499189
- DOI: 10.1534/genetics.116.198671
GSK-3β Homolog Rim11 and the Histone Deacetylase Complex Ume6-Sin3-Rpd3 Are Involved in Replication Stress Response Caused by Defects in Dna2
Abstract
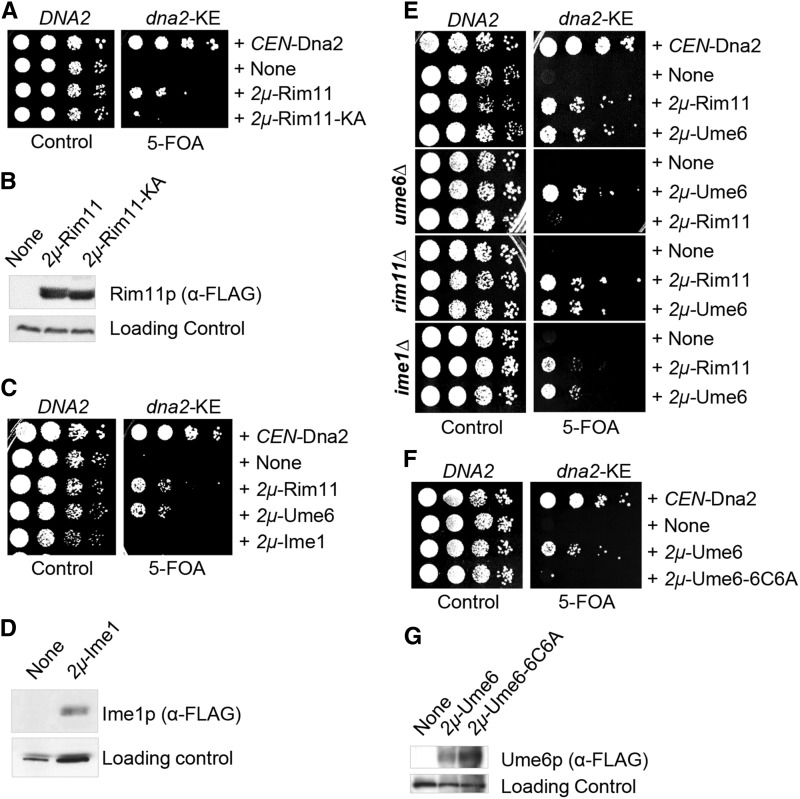
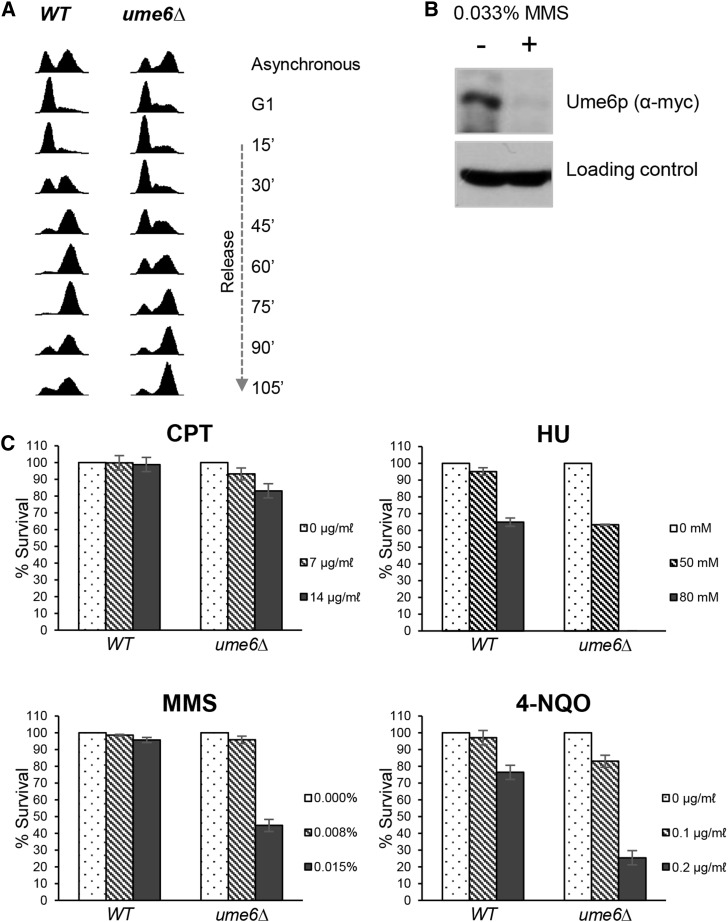
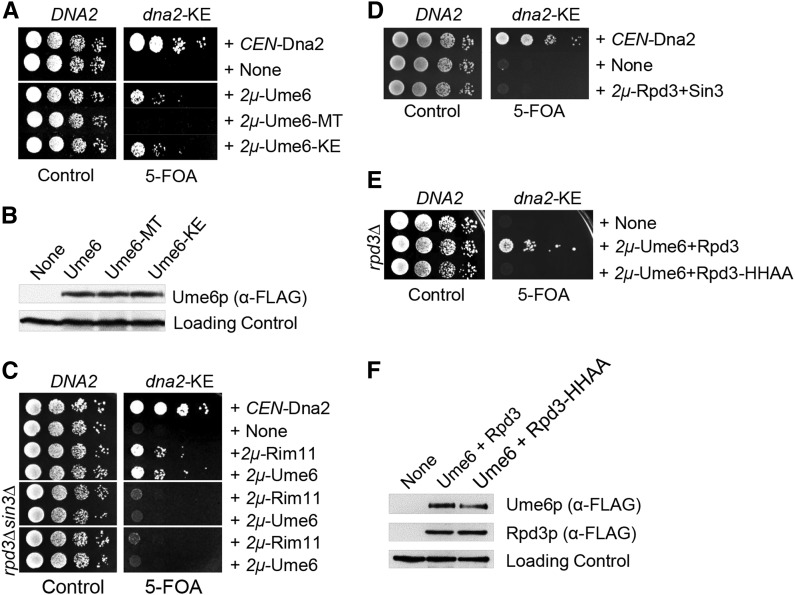
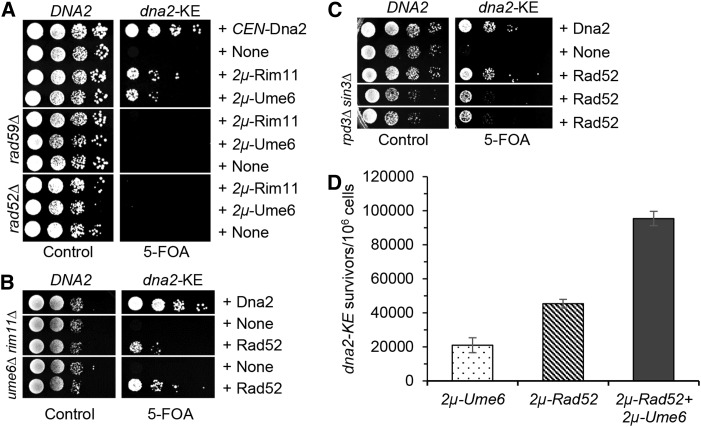
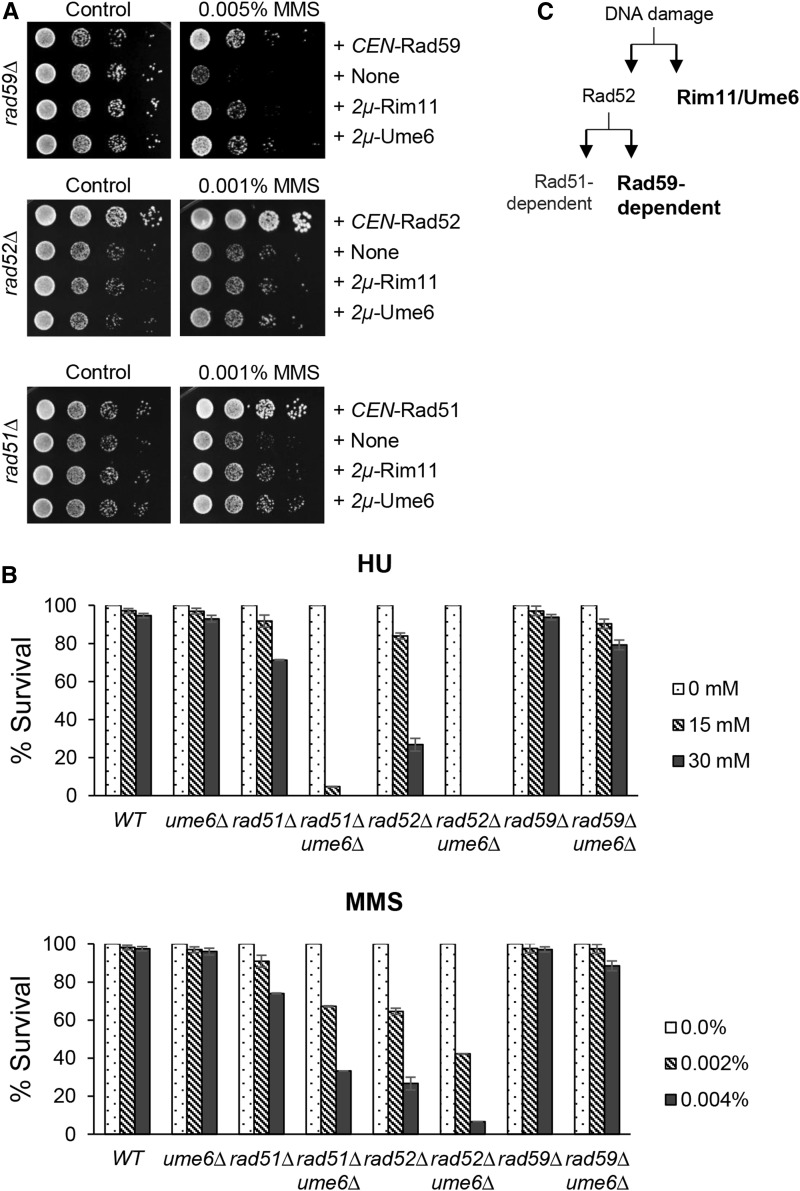
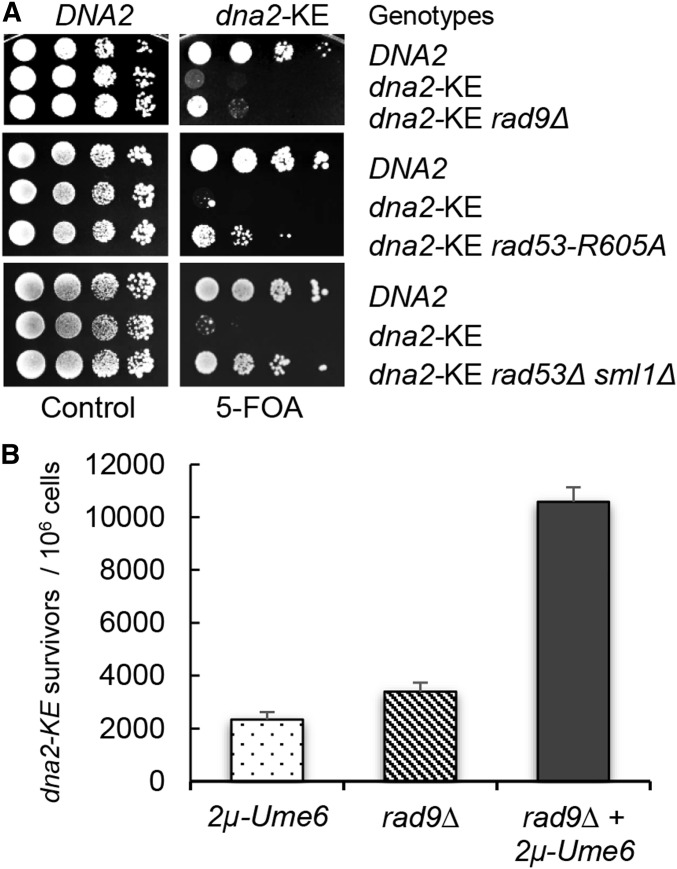
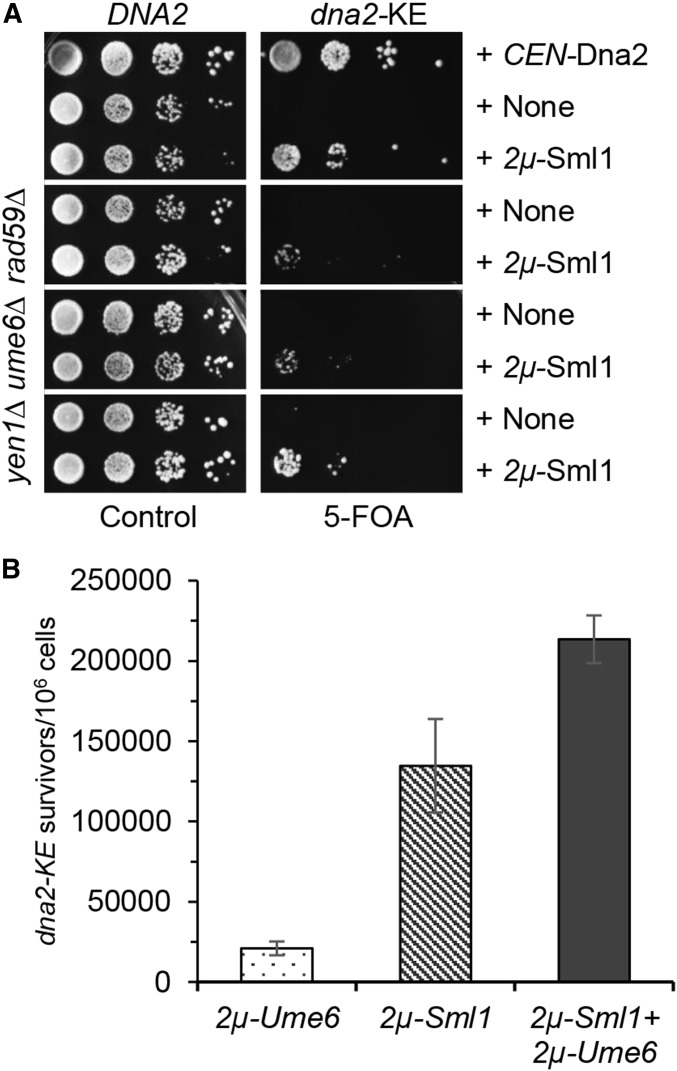
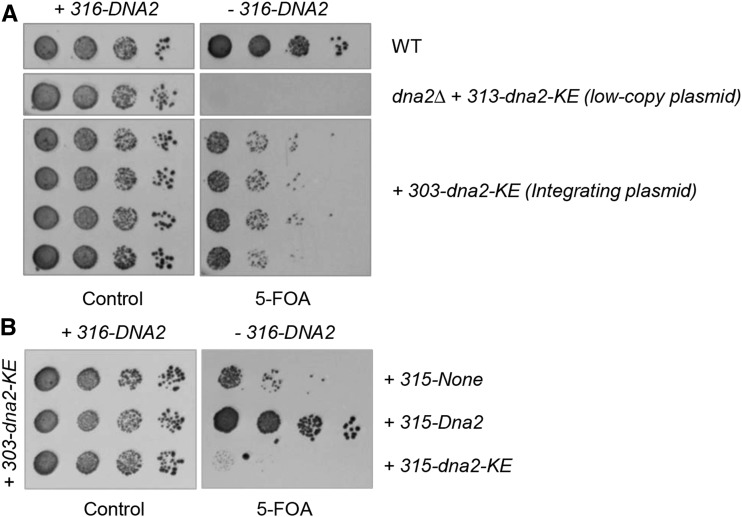
Lagging strand synthesis is mechanistically far more complicated than leading strand synthesis because it involves multistep processes and requires considerably more enzymes and protein factors. Due to this complexity, multiple fail-safe factors are required to ensure successful replication of the lagging strand DNA. We attempted to identify novel factors that are required in the absence of the helicase activity of Dna2, an essential enzyme in Okazaki-fragment maturation. In this article, we identified Rim11, a GSK-3β-kinase homolog, as a multicopy suppressor of dna2 helicase-dead mutant (dna2-K1080E). Subsequent epistasis analysis revealed that Ume6 (a DNA binding protein, a downstream substrate of Rim11) also acted as a multicopy suppressor of the dna2 allele. We found that the interaction of Ume6 with the conserved histone deacetylase complex Sin3-Rpd3 and the catalytic activity of Rpd3 were indispensable for the observed suppression of the dna2 mutant. Moreover, multicopy suppression by Rim11/Ume6 requires the presence of sister-chromatid recombination mediated by Rad52/Rad59 proteins, but not vice versa. Interestingly, the overexpression of Rim11 or Ume6 also suppressed the MMS sensitivity of rad59Δ. We also showed that the lethality of dna2 helicase-dead mutant was attributed to checkpoint activation and that decreased levels of deoxynucleotide triphosphates (dNTPs) by overexpressing Sml1 (an inhibitor of ribonucleotide reductase) rescued the dna2 mutant. We also present evidence that indicates Rim11/Ume6 works independently but in parallel with that of checkpoint inhibition, dNTP regulation, and sister-chromatid recombination. In conclusion, our results establish Rim11, Ume6, the histone deacetylase complex Sin3-Rpd3 and Sml1 as new factors important in the events of faulty lagging strand synthesis.
Keywords: Dna2; GSK-3b; HDAC; checkpoint; genome instability.
Copyright © 2017 by the Genetics Society of America.
Figures

References
-
- Bae S. H., Seo Y. S., 2000. Characterization of the enzymatic properties of the yeast dna2 Helicase/endonuclease suggests a new model for Okazaki fragment processing. J. Biol. Chem. 275: 38022–38031. - PubMed
-
- Bae S. H., Bae K. H., Kim J. A., Seo Y. S., 2001a RPA governs endonuclease switching during processing of Okazaki fragments in eukaryotes. Nature 412: 456–461. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
