

The active force-length relationship is invisible during extensive eccentric contractions in skinned skeletal muscle fibres
- PMID: 28469023
- PMCID: PMC5443931
- DOI: 10.1098/rspb.2016.2497
The active force-length relationship is invisible during extensive eccentric contractions in skinned skeletal muscle fibres
Abstract
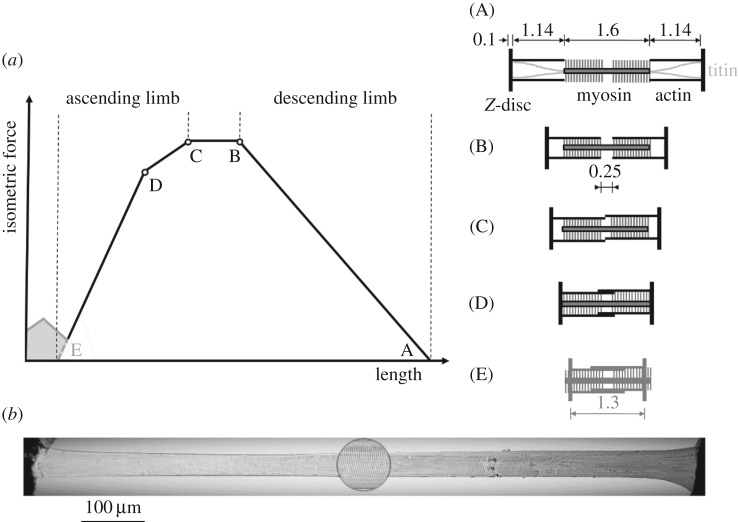
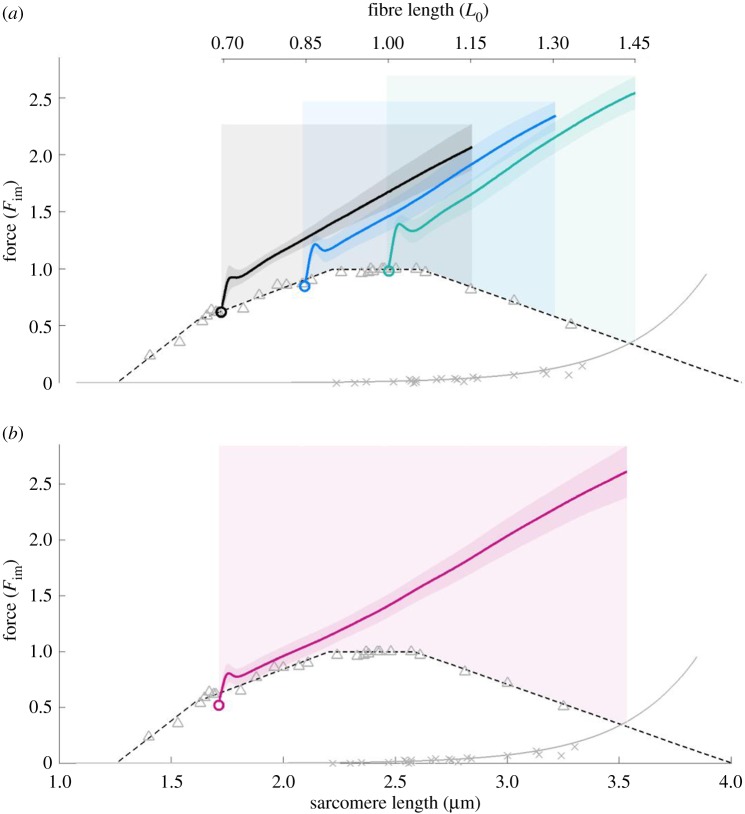
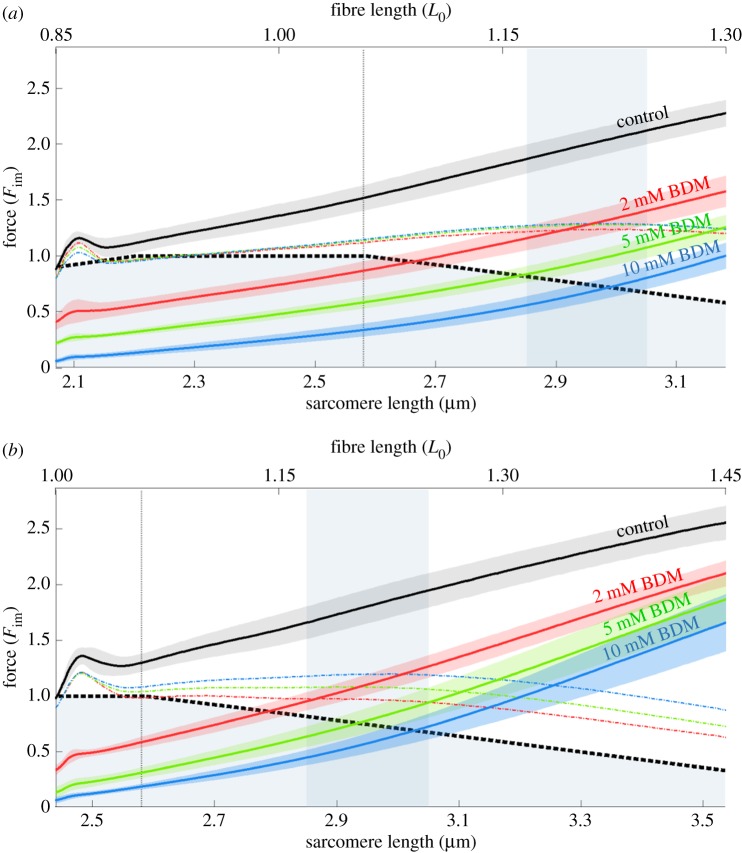
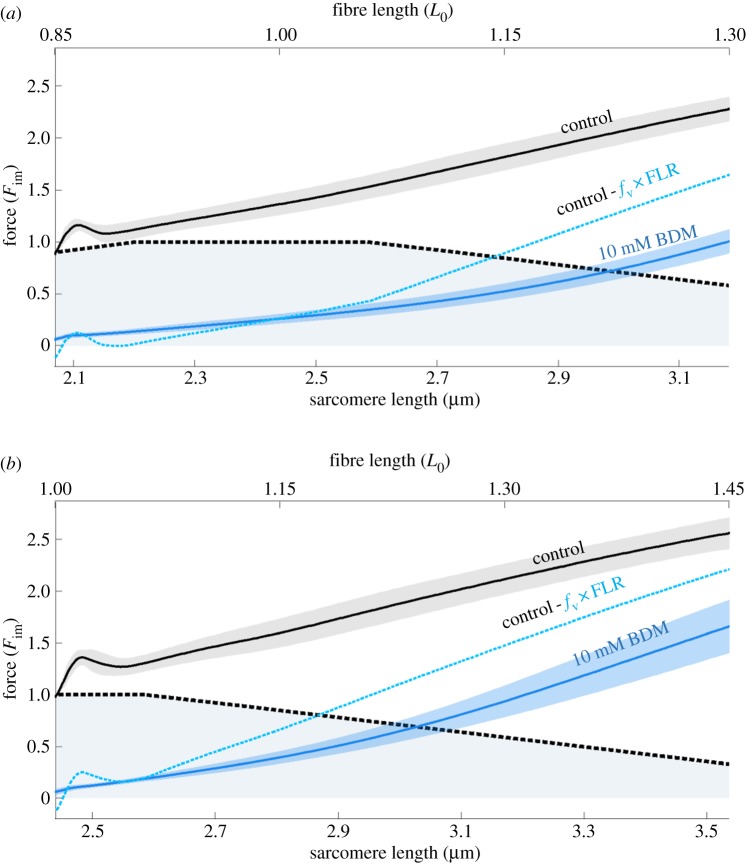
In contrast to experimentally observed progressive forces in eccentric contractions, cross-bridge and sliding-filament theories of muscle contraction predict that varying myofilament overlap will lead to increases and decreases in active force during eccentric contractions. Non-cross-bridge contributions potentially explain the progressive total forces. However, it is not clear whether underlying abrupt changes in the slope of the nonlinear force-length relationship are visible in long isokinetic stretches, and in which proportion cross-bridges and non-cross-bridges contribute to muscle force. Here, we show that maximally activated single skinned rat muscle fibres behave (almost across the entire working range) like linear springs. The force slope is about three times the maximum isometric force per optimal length. Cross-bridge and non-cross-bridge contributions to the muscle force were investigated using an actomyosin inhibitor. The experiments revealed a nonlinear progressive contribution of non-cross-bridge forces and suggest a nonlinear cross-bridge contribution similar to the active force-length relationship (though with increased optimal length and maximum isometric force). The linear muscle behaviour might significantly reduce the control effort. Moreover, the observed slight increase in slope with initial length is in accordance with current models attributing the non-cross-bridge force to titin.
Keywords: cross-bridge; lengthening contractions; linear muscle behaviour; muscle stretch; titin.
© 2017 The Author(s).
Conflict of interest statement
We declare we have no competing interests.
Figures
Similar articles
-
How velocity impacts eccentric force generation of fully activated skinned skeletal muscle fibers in long stretches.J Appl Physiol (1985). 2022 Jul 1;133(1):223-233. doi: 10.1152/japplphysiol.00735.2021. Epub 2022 Jun 2. J Appl Physiol (1985). 2022. PMID: 35652830
-
Energy cost of isometric force production after active shortening in skinned muscle fibres.J Exp Biol. 2017 Apr 15;220(Pt 8):1509-1515. doi: 10.1242/jeb.117622. Epub 2017 Feb 23. J Exp Biol. 2017. PMID: 28232399
-
Extensive eccentric contractions in intact cardiac trabeculae: revealing compelling differences in contractile behaviour compared to skeletal muscles.Proc Biol Sci. 2019 May 29;286(1903):20190719. doi: 10.1098/rspb.2019.0719. Epub 2019 May 29. Proc Biol Sci. 2019. PMID: 31138072 Free PMC article.
-
Mechanisms of enhanced force production in lengthening (eccentric) muscle contractions.J Appl Physiol (1985). 2014 Jun 1;116(11):1407-17. doi: 10.1152/japplphysiol.00069.2013. Epub 2013 Feb 21. J Appl Physiol (1985). 2014. PMID: 23429875 Review.
-
Residual Force Enhancement Following Eccentric Contractions: A New Mechanism Involving Titin.Physiology (Bethesda). 2016 Jul;31(4):300-12. doi: 10.1152/physiol.00049.2014. Physiology (Bethesda). 2016. PMID: 27252165 Review.
Cited by
-
A continuum-mechanical skeletal muscle model including actin-titin interaction predicts stable contractions on the descending limb of the force-length relation.PLoS Comput Biol. 2017 Oct 2;13(10):e1005773. doi: 10.1371/journal.pcbi.1005773. eCollection 2017 Oct. PLoS Comput Biol. 2017. PMID: 28968385 Free PMC article.
-
Impact of lengthening velocity on the generation of eccentric force by slow-twitch muscle fibers in long stretches.Pflugers Arch. 2024 Oct;476(10):1517-1527. doi: 10.1007/s00424-024-02991-4. Epub 2024 Jul 24. Pflugers Arch. 2024. PMID: 39043889 Free PMC article.
-
Fast stretching of skeletal muscle fibres abolishes residual force enhancement.J Exp Biol. 2022 May 15;225(10):jeb244011. doi: 10.1242/jeb.244011. Epub 2022 May 30. J Exp Biol. 2022. PMID: 35485194 Free PMC article.
-
Cross-Bridges and Sarcomeric Non-cross-bridge Structures Contribute to Increased Work in Stretch-Shortening Cycles.Front Physiol. 2020 Jul 28;11:921. doi: 10.3389/fphys.2020.00921. eCollection 2020. Front Physiol. 2020. PMID: 32848862 Free PMC article.
-
Residual force enhancement is not altered while force depression is amplified at the cellular level in old age.J Exp Biol. 2025 Jan 1;228(1):JEB248155. doi: 10.1242/jeb.248155. Epub 2025 Jan 13. J Exp Biol. 2025. PMID: 39688124 Free PMC article.
References
-
- Hill AV. 1938. The heat of shortening and the dynamic constants of muscle. Proc. R. Soc. Lond. B 126, 136–195. (10.1098/rspb.1938.0050) - DOI
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
