

A HAND to TBX5 Explains the Link Between Thalidomide and Cardiac Diseases
- PMID: 28469241
- PMCID: PMC5431093
- DOI: 10.1038/s41598-017-01641-3
A HAND to TBX5 Explains the Link Between Thalidomide and Cardiac Diseases
Abstract
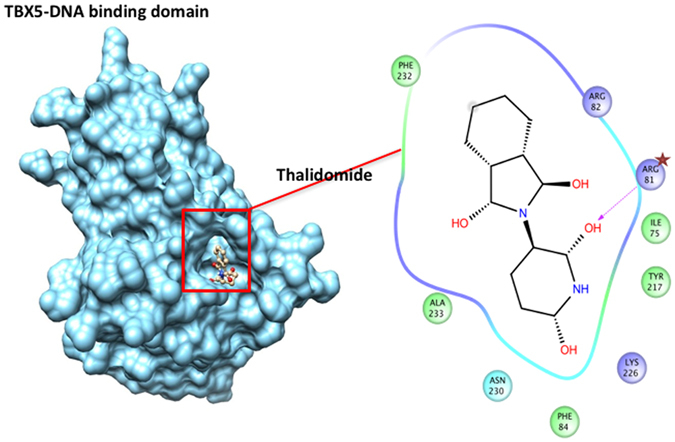
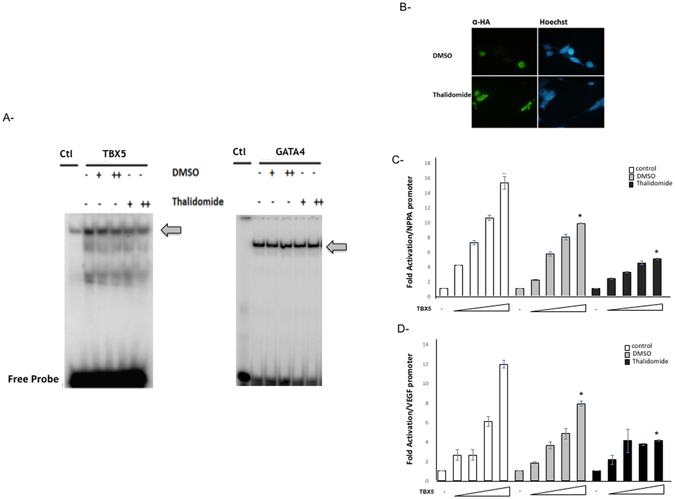
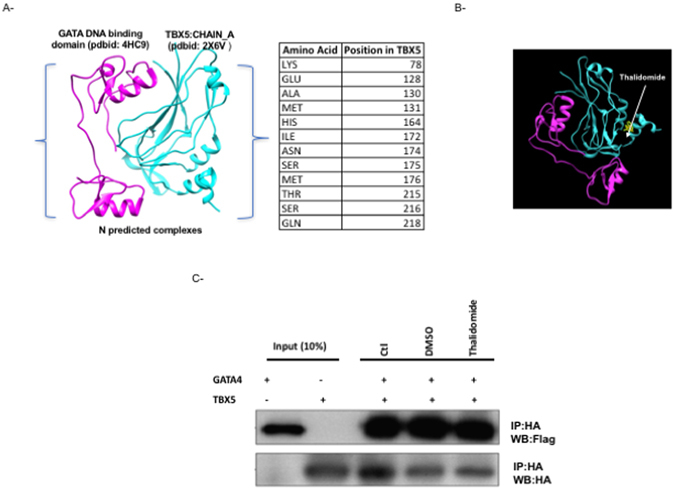
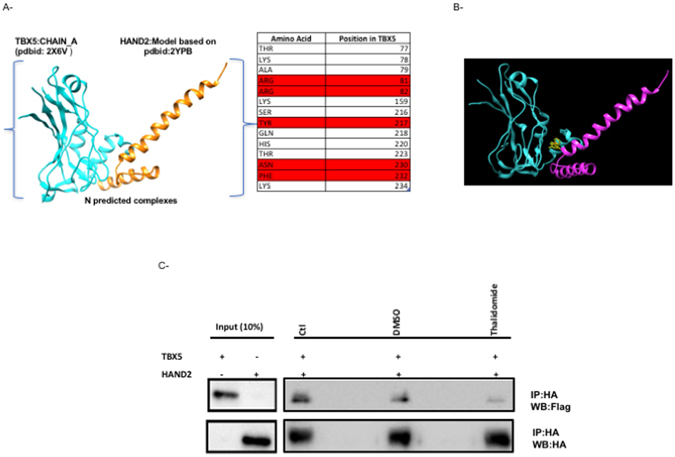
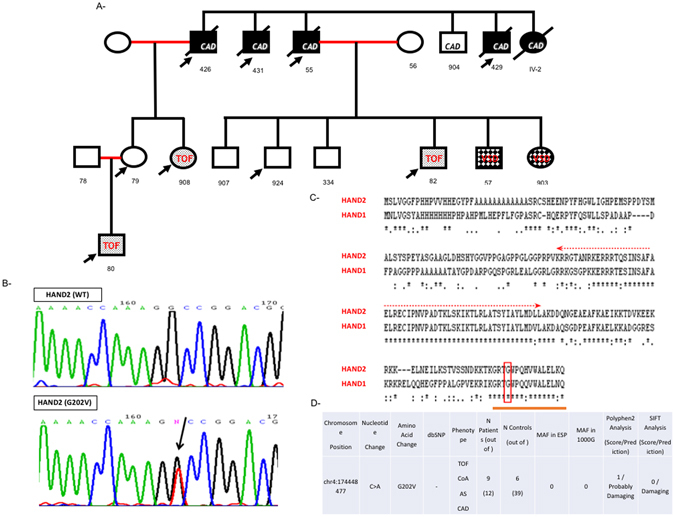
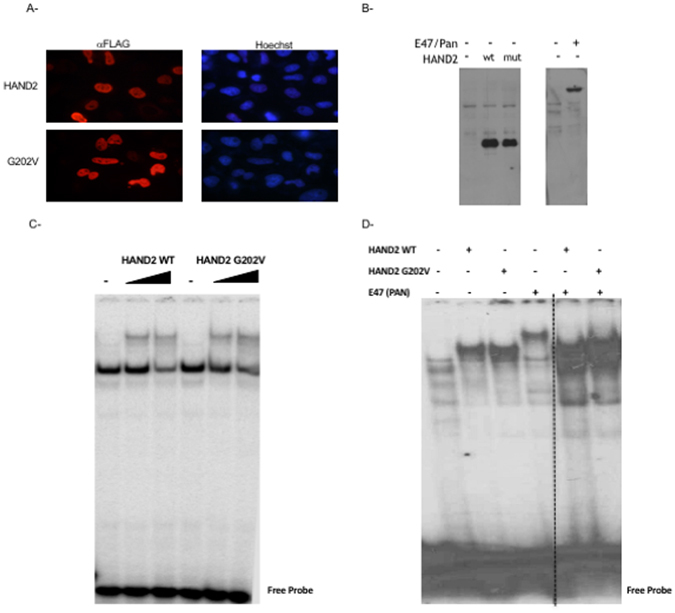
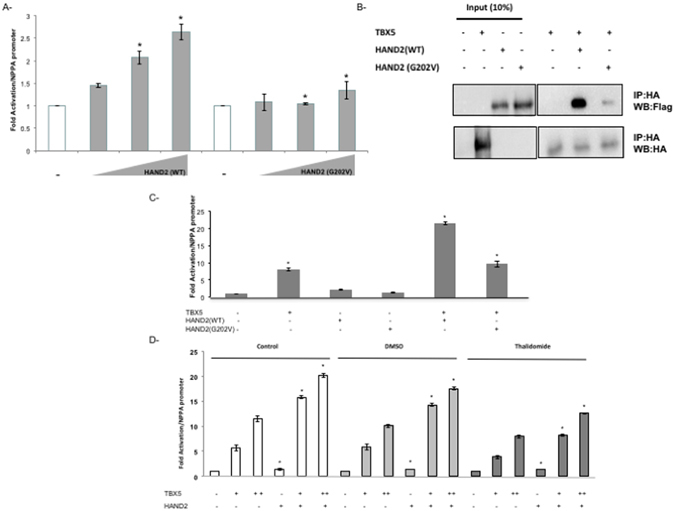
Congenital heart disease is the leading cause of death in the first year of life. Mutations only in few genes have been linked to some cases of CHD. Thalidomide was used by pregnant women for morning sickness but was removed from the market because it caused severe malformations including CHDs. We used both in silico docking software, and in vitro molecular and biochemical methods to document a novel interaction involving Thalidomide, TBX5, and HAND2. Thalidomide binds readily to TBX5 through amino acids R81, R82, and K226 all implicated in DNA binding. It reduces TBX5 binding to DNA by 40%, and suppresses TBX5 mediated activation of the NPPA and VEGF promoters by 70%. We documented a novel interaction between TBX5 and HAND2, and showed that a p.G202V HAND2 variant associated with CHD and coronary artery diseases found in a large Lebanese family with high consanguinity, drastically inhibited this interaction by 90%. Similarly, thalidomide inhibited the TBX5/HAND2 physical interaction, and the in silico docking revealed that the same amino acids involved in the interaction of TBX5 with DNA are also involved in its binding to HAND2. Our results establish a HAND2/TBX5 pathway implicated in heart development and diseases.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
Possible New Candidates Involved to Thalidomide-Related Limbs and Cardiac Defects: A Systems Biology Approach.Biochem Genet. 2025 Jun;63(3):2140-2160. doi: 10.1007/s10528-024-10790-w. Epub 2024 Apr 30. Biochem Genet. 2025. PMID: 38689186
-
Genetic evaluation of HAND2 gene and its effects on thalidomide embryopathy.Birth Defects Res. 2022 Dec 1;114(20):1354-1363. doi: 10.1002/bdr2.2092. Epub 2022 Sep 30. Birth Defects Res. 2022. PMID: 36177858
-
Structural basis of TBX5-DNA recognition: the T-box domain in its DNA-bound and -unbound form.J Mol Biol. 2010 Jul 2;400(1):71-81. doi: 10.1016/j.jmb.2010.04.052. Epub 2010 May 5. J Mol Biol. 2010. PMID: 20450920
-
Congenital heart diseases and their association with the variant distribution features on susceptibility genes.Clin Genet. 2017 Mar;91(3):349-354. doi: 10.1111/cge.12835. Epub 2016 Sep 5. Clin Genet. 2017. PMID: 27426723 Review.
-
Hand factors as regulators of cardiac morphogenesis and implications for congenital heart defects.Birth Defects Res A Clin Mol Teratol. 2011 Jun;91(6):485-94. doi: 10.1002/bdra.20796. Epub 2011 Apr 1. Birth Defects Res A Clin Mol Teratol. 2011. PMID: 21462297 Free PMC article. Review.
Cited by
-
Epigenetics and Congenital Heart Diseases.J Cardiovasc Dev Dis. 2022 Jun 9;9(6):185. doi: 10.3390/jcdd9060185. J Cardiovasc Dev Dis. 2022. PMID: 35735814 Free PMC article. Review.
-
Possible New Candidates Involved to Thalidomide-Related Limbs and Cardiac Defects: A Systems Biology Approach.Biochem Genet. 2025 Jun;63(3):2140-2160. doi: 10.1007/s10528-024-10790-w. Epub 2024 Apr 30. Biochem Genet. 2025. PMID: 38689186
-
Transcriptomic Alterations in Lung Adenocarcinoma Unveil New Mechanisms Targeted by the TBX2 Subfamily of Tumor Suppressor Genes.Front Oncol. 2018 Oct 30;8:482. doi: 10.3389/fonc.2018.00482. eCollection 2018. Front Oncol. 2018. PMID: 30425966 Free PMC article.
-
Single-Construct Polycistronic Doxycycline-Inducible Vectors Improve Direct Cardiac Reprogramming and Can Be Used to Identify the Critical Timing of Transgene Expression.Int J Mol Sci. 2017 Aug 19;18(8):1805. doi: 10.3390/ijms18081805. Int J Mol Sci. 2017. PMID: 28825623 Free PMC article.
-
Thalidomide remodels developing heart in chick embryo: discovery of a thalidomide mediated hematoma in heart muscle.Naunyn Schmiedebergs Arch Pharmacol. 2018 Oct;391(10):1093-1105. doi: 10.1007/s00210-018-1532-2. Epub 2018 Jul 7. Naunyn Schmiedebergs Arch Pharmacol. 2018. PMID: 29982937
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
